King’s amino-acid findability argument
Whereas Kimura (1968) proposed his version of the Neutral Theory of Molecular Evolution as the answer to an esoteric problem of population genetics theory, King and Jukes (1969) proposed a theory driven by the results of macromolecular sequence comparisons. Molecular evolution, in their view, demanded “new rules.” As evidence for neutrality, they pointed to a general correspondence between the frequencies of amino acids in protein sequences, and the frequencies expected from translating randomly generated sequences with the genetic code.
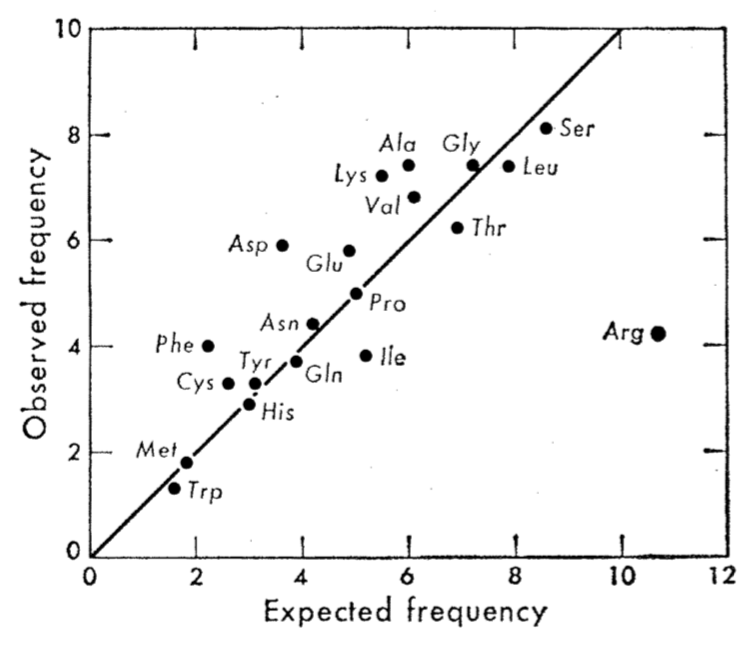
At the bottom left are Met and Trp with 1 codon each and then, proceeding with some variation up and to the right, we have the 2-codon blocks (Cys, His, Tyr, Phe, Gln, Asn, Asp, Lys), Ile with 3 codons, then the 4-codon blocks (Pro, thr, Val, Ala, Gly) then the 6-codon blocks (Leu, Ser, Arg), with Arg being a somewhat extreme outlier due to the CpG effect.
However, King rather quickly recanted on this argument. It is rare for a scientist to do that, so pay attention. In the proceedings of a 1971 conference that came out 2 years later, King (1973) said that this was not evidence of neutrality, but rather evidence for some kind of indeterministic process dependent on mutation. He explains
“If a gene is in the process of progressive, adaptive evolution, there might very likely be more than one among the thousand or so possible single-step changes that would be evolutionarily advantageous. Then the first of these to occur by mutation would have the first chance to take over. The conditions of selection would then be changed, and it would be too late for the other previously potential candidates. Thus the probability of fixation [probability of origin-and-fixation] of an amino acid is a function of its frequency of arising by mutation, and this will happen more often to amino acids with more codons. The eventual distribution of amino acid frequencies will reflect, more or less passively, the peculiarities of an arbitrary genetic code, even if most evolutionary changes are due exclusively to Darwinian adaptive evolution.” (p. 7)
King JL. 1973. The Influence of the Genetic Code on Protein Evolution. In: Schoffeniels E, editor. Biochemical Evolution and the Origin of Life. Viers: North-Holland Publishing Company. p. 3-13
King did not generalize further on this argument. However, we can see this as an instance of a general form of argument in which phenotypes that are over-represented in genotype-space are more findable due to mutation. Amino acids with larger numbers of codons (in the genetic code) occupy a greater volume of sequence space, analogous to phenotypes with large numbers of genotypes in genotype-space, and this makes them more findable by an evolutionary process that explores sequence space (or genotype space) via mutations. So the amino acids with the most codons will tend to be the most common in proteins, and this argument does not require neutrality, but merely a process subject to biases in introduction of amino acid phenotypes.
The form of this argument is analogous to that made by Ard Louis and colleagues in regard to RNA folds that are common in sequence space (Dingle, et al. 2022).
King’s two conference papers in this period reveal important thinking about evolution in discrete spaces. King (1973) gives this image combining states, paths, upwardness, and a fitness landscape. Note that this is not merely a passive depiction of “Maynard Smith’s” concept, but represents creative and synthetic thinking.
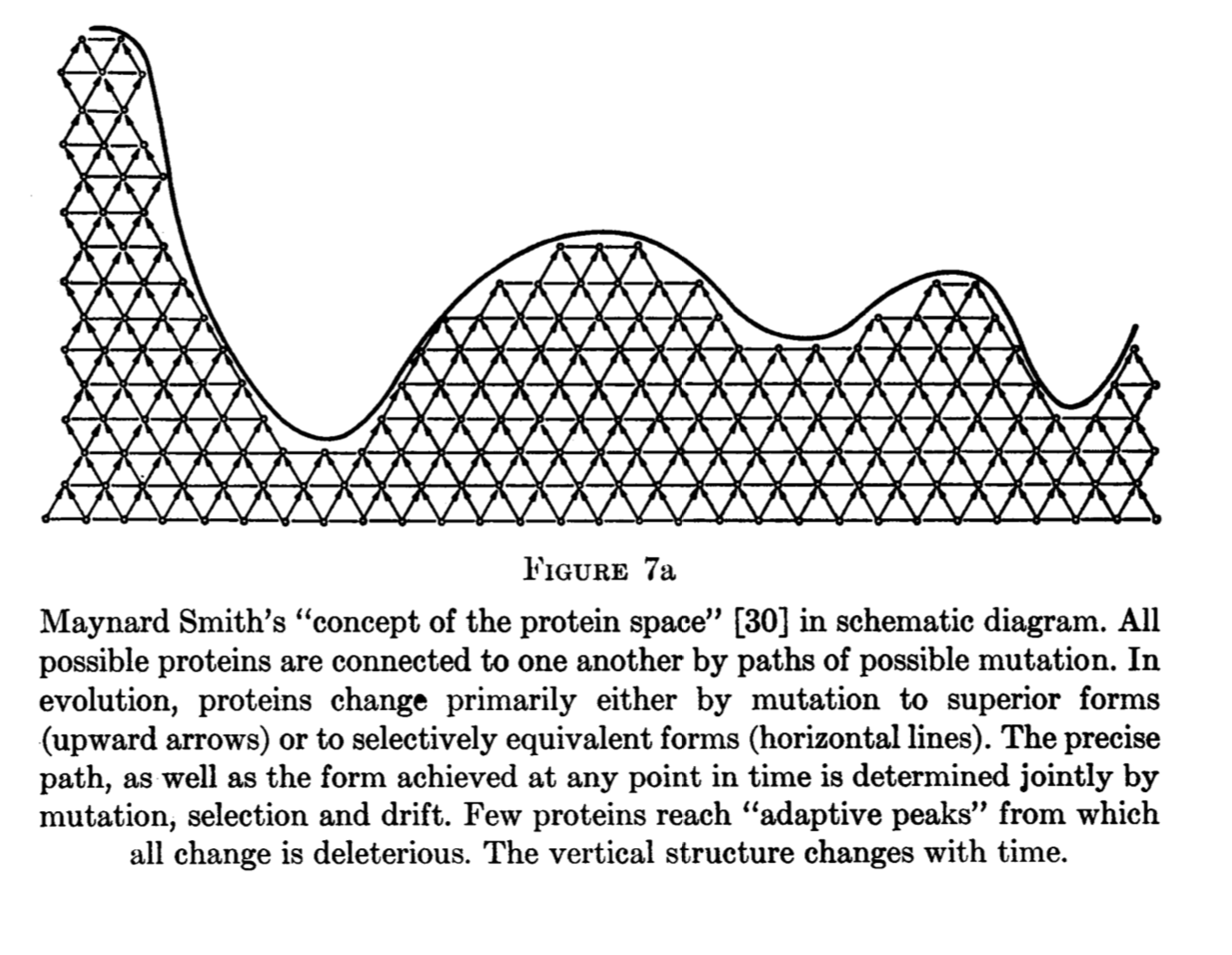
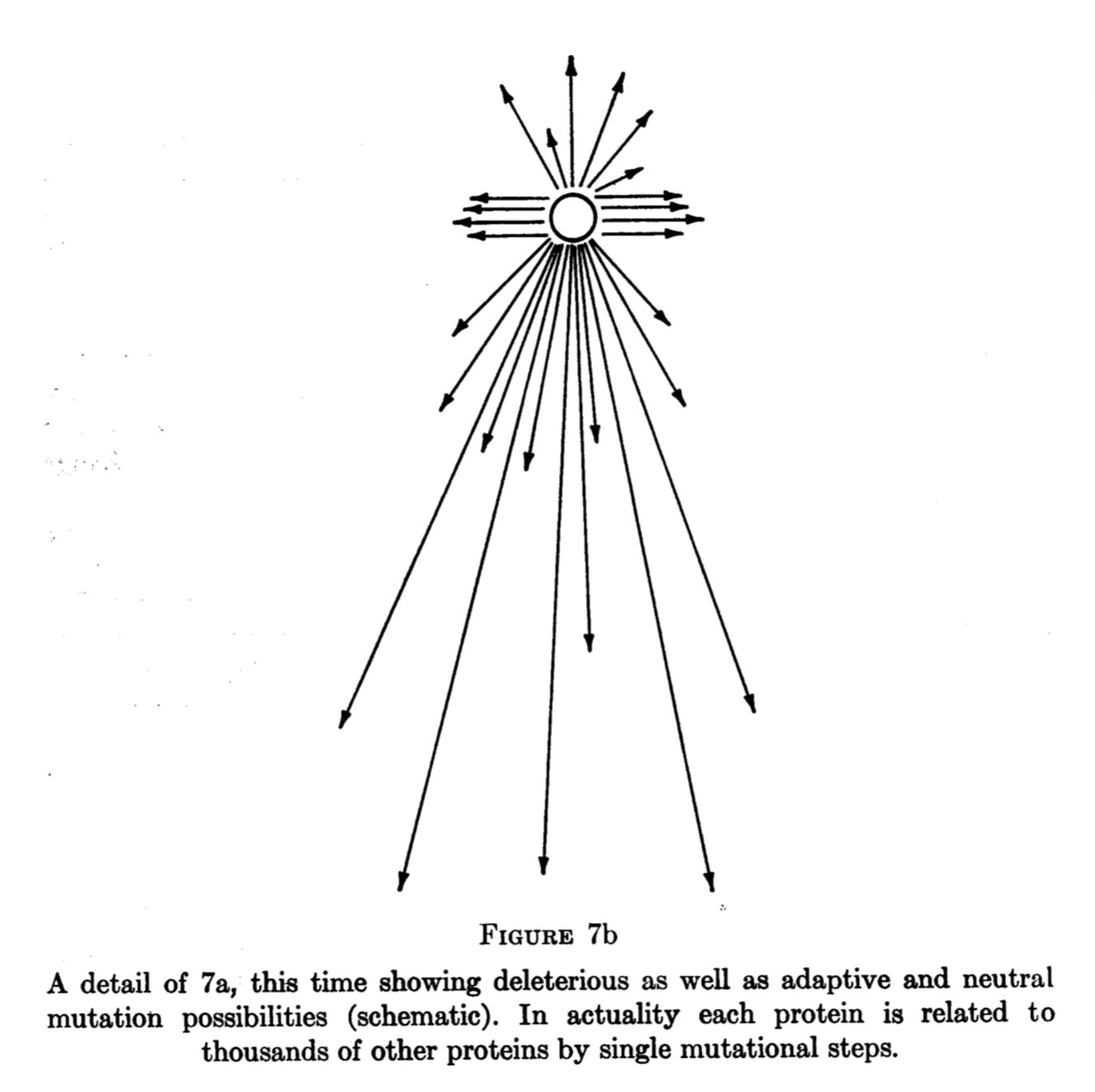
What happened to King’s argument? It could have, but apparently did not, stimulate a model like that of Yampolsky and Stoltzfus (2001). However, it was not completely lost to history. Leigh Van Valen repeated the argument in a 1974 paper, attributing it ambiguously either to an “oral” clarification of Lewontin or to Stebbins and Lewontin (1973) :
“2. Lewontin also made more plausible another rebuttal that Stebbins and Lewontin (1973) made to a neutralist argument. The latter argument is the general similarity of the proportions of amino acids in proteins to the proportions of their respective codons among all codons, given the observed proportions of the four nucleotides. Now consider a protein sitting in the protein space. There may be several sequences (local adaptive peaks) better than the one it now has. However, these will be unequally available. The most available will be those for which the needed mutations are for amino acids with the most codons, assuming that many of the possible steps to the peak increase fitness. Once the protein has chosen its peak, the final sequence is determined only by selection. Therefore selection can give a correspondence of proportions as easily as drift can.”
The article from Stebbins and Lewontin appears in the same symposium volume as King’s paper. So, all 3 of them probably went to the same 1971 meeting and were exposed to this idea (the papers were not published until 1973). However, the version of the argument in Stebbins and Lewontin is inadequate and cannot have been the source of Van Valen’s version, which is as clear as that of King. So, either Van Valen got it from King but misattributed it to Lewontin, or Lewontin’s “oral” clarification included a greatly improved argument.
Maynard Smith (1975) also repeats King’s argument in his comments on the neutralist-selectionist controvery, concluding that:
“Hence the correlation does not enable us to decide between the two. However, it is worth remembering that if we accept the selectionist view that most substitutions are selective, we cannot at the same time assume that there is a unique deterministic course for evolution. Instead, we must assume that there are alternative ways in which a protein can evolve, the actual path taken depending on chance events. This seems to be the minimum concession the selectionists will have to make to the neutralists; they may have to concede much more.”
I love this quotation because it reveals a world that most people do not know existed in 1975, a world in which selectionists were not yet entertaining the idea of evolution as a Markov chain of origin-fixation events, but were still using the shifting-gene-frequencies view defended by Stebbins and Lewontin (1973).
References
King JL. 1973. The Role of Mutation in Evolution. In: Sixth Berkeley Symposium on Mathematical Statistics and Probability (eds Le Cam, Neyman, and Scott) Berkeley, California.
King JL. 1971. The Influence of the Genetic Code on Protein Evolution. In: Schoffeniels E, editor. Biochemical Evolution and the Origin of Life. Viers: North-Holland Publishing Company. p. 3-13.
Maynard Smith J. 1975. The Theory of Evolution. Cambridge: Cambridge University Press.
Stebbins GL, Lewontin RC, 1973. Comparative evolution at the levels of molecules, organisms and populationseditors. In: Sixth Berkeley Symposium on Mathematical Statistics and Probability (eds Le Cam, Neyman, and Scott) Berkeley, California
Van Valen L. 1974. Molecular evolution as predicted by natural selection. Journal of Molecular Evolution 3:89-101.