In a recent QRB paper with David McCandlish, we review the form, origins, uses, and implications of models (e.g., the familiar K = 4Nus) that represent evolutionary change as a 2-step process of (1) the introduction of a new allele by mutation, followed by (2) its fixation or loss.
What could be surprising about these “origin-fixation” models, which are invoked in theoretical models of adaptation (e.g., the mutational landscape model) and in widely used methods applied to phylogenetic inference, comparative genomics, detecting selection, modeling codon usage, and so on?
Quite a lot, it turns out. For one thing, there are 2 distinct flavors— aggregate-rate and sequential fixation. They look similar in some cases, but they mean different things. It was David who realized this, whereas in past work, I had not distinguished the two (e.g., in my NK model of protein adaptation, I implemented a sequential fixation model while citing a reference for an aggregate-rate model).
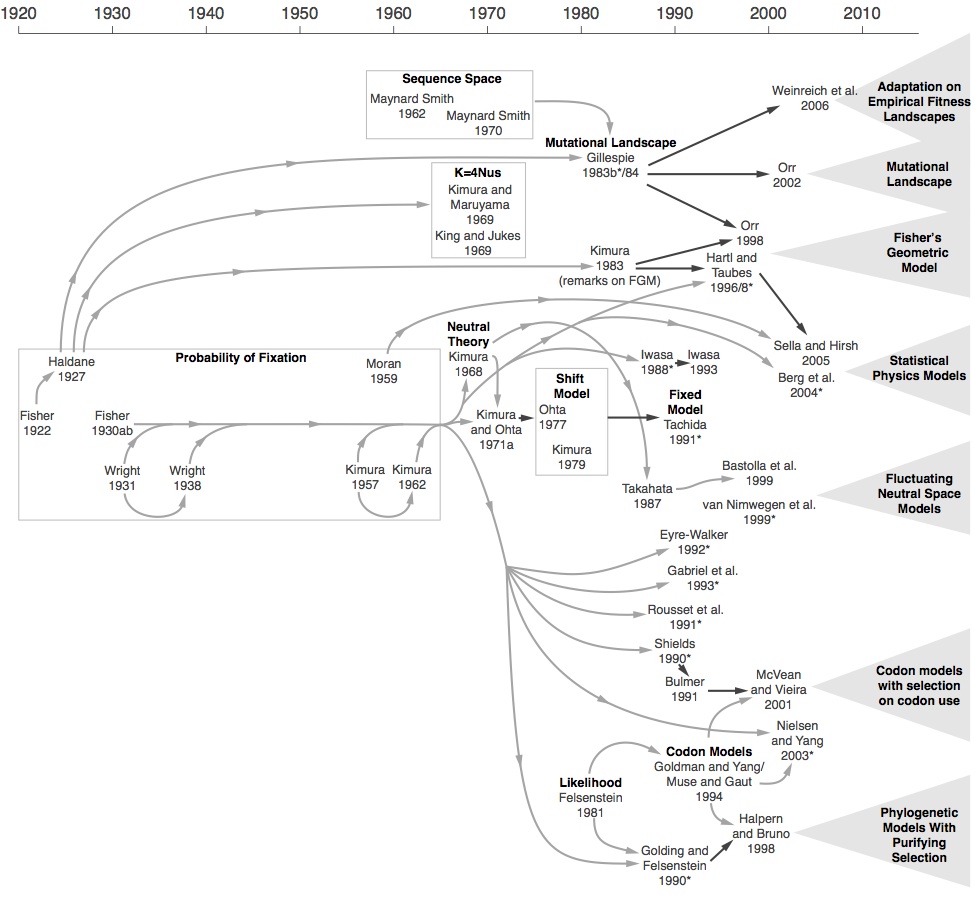
But there are bigger surprises. In spite of being simple in form and widely used, aggregate-rate models did not appear until 1969 (by King & Jukes, and Kimura & Maruyama), in the midst of the molecular revolution; the first sequential fixation model is from Gillespie (1983), but these models were re-invented several times independently, as shown in the figure below.
Haldane, Fisher and Wright knew about the probability of fixation of a new mutation starting in the 1920s. Why didn’t they invent origin-fixation models? The answer is yet another surprise: they just did not believe that evolution occurs by a 2-step process of mutation followed by fixation or loss. They had a more complicated “gene pool” view in which evolution is a shift in the relative frequencies of alleles that are already present in the population, without new mutations.
Think about it like this. Evolutionary genetics, as we understand it today, covers a lot of ground and allows many different modes of evolution. Origin-fixation models invoke a regime of evolutionary genetics in which the power of mutation to determine the rate and direction of evolution is maximized, recalling the mutationist theory of early geneticists; in sharp contrast, the Modern Synthesis invoked a regime in which the power of mutation is extremely weak and the power of selection is strong.
And the final surprise is how we (the evolutionary genetics community) have been addressing this epic clash of world-views: for the most part, we’ve been ignoring it. You would think this would be burning issue #1 for evolutionary geneticists. You would think educators and science journalists would have heard the news that the fabled architects have suffered a scientific loss at the hands of the much-maligned mutationists. But there has been no debate. Instead, as we write, “origin-fixation models are frequently invoked without regard to either their underlying assumptions or their implications, as if they represented a generic framework for modeling the evolutionary process.” One of the aims of our review is to draw attention to origin-fixation models as a distinctive theory of evolution to be tested, rather than merely a method to be applied uncritically.

Leave a comment