The Mutationism Myth (3): Foundations of evolutionary genetics
This is the third in a series of 2010 blogs entitled “The Mutationism Myth” (a more scholarly version of this material ended being published in J. Hist. Biol. by Stoltzfus and Cable, 2014)
In this oft-told story (see part 1), the discovery of genetics in 1900 leads to rejection of Darwin’s theory and the rise of “mutationism”, a laughable1 theory that imagines evolution by mutation alone, without selection. “Mutationism” prevails for a generation, until Fisher, Haldane and Wright show that genetics is the missing key to Darwinism. In the conclusion to the story, the world is set right again when the “Modern Synthesis”, combining selection with Mendelian genetics, shoulders aside the mutationist heresy, which ends up in the dustbin of history with the other “doomed rivals” of Darwin’s great theory.2
Thats the story, at least. In reality— as we found out in part 2—, the Mendelians rejected Darwin’s errant principles of heredity, not his principle of selection. What kind of view did the Mendelians develop? Addressing this question is our next challenge. Today, in part 3, we’ll consider aspects of the Mendelian view that became the foundations of mainstream 20th-century thinking. In part 4, we’ll delve into some “non-Darwinian” or “anti-Darwinian” aspects that were rejected, or merely ignored.
The Mutationism Myth. 3. Foundations of evolutionary genetics
Darwin’s “Natural Selection” theory posited a smooth and automatic process of adaptation to altered conditions, dependent on infinitesimal hereditary fluctuations (“indefinite variability”, in Darwin’s terminology) induced by the effect of “altered conditions of life” on the “sexual organs”. As we discovered in part 2, geneticists rejected fluctuation because it is incompatible with the assumption of exclusively Mendelian inheritance, an assumption embraced eagerly by geneticists, and held in suspicion by others for many years. As Bateson wrote:
“To Darwin the question, What is a variation? presented no difficulties. Any difference between parent and offspring was a variation. Now we have to be more precise. First we must, as de Vries has shown, distinguish real, genetic, variation from fluctuational variations, due to environmental and other accidents, which cannot be transmitted.” (p. 95)
and as Morgan wrote:
“As has been explained, the kind of variability on which Darwin based his theory of natural selection can no longer be used in support of that theory, because, in the first place, in so far as fluctuating variations are due to environmental effect, these differences are now known not to be inherited, and because, in the second place, selection of the differences between individuals, due to the then existing genetic variants, while changing the number of individuals of a given kind, will not introduce anything new. The essential [feature] of the evolutionary process is the occurrence of new characteristics.” p. 148-149 of Morgan (1932) 3
Because heredity and variation did not behave in the manner assumed by Darwin and his followers, it was up to a new generation of evolutionists to develop a new understanding of evolution. Thus, at a time when naturalists were dismissing genetics and clinging to 19th-century views of heredity, including Darwinism and Lamarckism, a group of Young Turks4 was laying the foundations of the genetics-based understanding of evolution that dominated the 20th century.
The concept of population genetics
To understand these foundations, I need to say a few words about the theoretical side of evolutionary genetics, often referred to as “population genetics”. Please recall from Theory vs. Theory that when we talk about population genetics theory or music theory, thats a different sense of “theory” from Lamarck’s theory or the prion theory of disease. Previously, we called them theoryA (body of abstract principles) and theoryC (grand conjecture).
Population genetics theoryA (roughly speaking) works out the implications of transmission genetics in populations of reproducing organisms, focusing on implications of such Mendelian phenomena as biparental inheritance, chromosome assortment, mutation, recombination, sex-linked inheritance, and so on.
As it exists today, population genetics theoryA covers a wide range of possible worlds, and thus a wide range of possible theoriesC. For instance, it provides classic equations to treat allele frequencies continuously and deterministically (e.g., Hardy-Weinberg), and at the same time, it provides another framework for addressing probabilistic changes with random drift. Is evolution deterministic or probabilistic? Population genetics theory2 doesn’t say- it allows us to consider both possibilities. Is evolutionary change smooth or does it come in chunks? Population genetics theoryA doesn’t say: it provides a quantitative genetics framework for continuous changes in quantitative characters, and a completely different framework for molecular evolutionists examining discrete characters. There are limiting cases where these different frameworks converge in some respects, but there is not any single realizable world in which all of population genetics theoryA applies, thus theoretical population genetics can’t be understood as a theoryC.
Crudely speaking, three frameworks of population genetics theory have been important in the 20th century: the stability analysis5 of systems of continuous allele frequencies, initially deterministic a la Hardy-Weinberg (or Lewontin-Kojima and so on) and later stochastic; the “quantitative genetics” theory of generational change in continuous-valued phenotypic characters (with implicit genetics) subject to selection; and the dynamics of the steady-state origin-fixation process, which was not an important paradigm until the neutral theory.
The Bateson-Saunders equilibrium
In a landmark 1902 report to the Evolution committee of the Royal Society, Bateson and Saunders report some of their own findings and, more generally, try to explain the new science of Mendelian genetics, and the implications of Mendel’s rules for evolution. In one of many fascinating comments, Bateson and Saunders suggeest that:
“It will be of great interest to study the statistics of such a population [with recognizable Mendelian characters] in nature. If the degree of dominance can be experimentally determined, or the heterozygote recognised, and we can suppose that all forms mate together with equal freedom and fertility, and that there is no natural selection in respect of the allelomorphs, it should be possible to predict the proportions of the several components of the population with some accuracy. Conversely, departures from the calculated result would then throw no little light on the influence of disturbing factors, selection, and the like.”
Those of you who know your population genetics will recognize, in this passage, a paradigm that continues to play a key role in contemporary research as a “zero-force” model, describing the case of an unperturbed system, i.e., a system at rest. Deviations from this resting state indicate the perturbing effect of some factor or force.
In 1908, Hardy and Weinberg independently derived solutions for the frequencies of genotypes and alleles in the zero-force model of Bateson and Saunders. The mathematical solution to the Hardy-Weinberg equilibrium, as it came to be called, is sufficiently trivial that publishing it was nearly beneath the dignity of G.H. Hardy, the archetypal pure mathematician. In his paper, Hardy seems to sneer at biologists, saying “I should have expected the very simple point which I wish to make to have been familiar to biologists”. Legend has it that Hardy learned of this problem while playing cricket with Punnett, the Mendelian, providing an early example of how interdisciplinary work is done.
The research program that eventually developed around this model was exactly as Bateson and Saunders imagined: compute the Hardy-Weinberg equilibrium, compare this to the observed frequencies, then interpret any deviations in terms of “the influence of disturbing factors”. Researchers continue to use it, as one may find by searching PubMed with “hardy-weinberg AND 2009 [date]“, which yields 532 publications for 2009. Contemporary philosophers discussing causation in evolutionary theory make frequent reference to Hardy-Weinberg as a zero-force law (see Stephens, 2001).
Given the crystal-clear statement of the problem by Bateson and Saunders, including the assumptions and the interpretive framework, should we not call it the Bateson-Saunders-Hardy-Weinberg equilibrium (or the Bateson-Saunders-Weinberg equilibrium, saving Hardy the embarrassment of receiving credit for something practical 6)?
Morgan’s origin-fixation process
An entirely different, but similarly prescient, model is found in T.H. Morgan‘s 1916 book:
“If through a mutation a character appears that is neither advantageous nor disadvantageous, but indifferent, the chance that it may become established in the race is extremely small, although by good luck such a thing may occur rarely. It makes no difference whether the character in question is a dominant or a recessive one, the chance of its becoming established is exactly the same. If through a mutation a character appears that has an injurious effect, however slight this may be, it has practically no chance of becoming established.
If through a mutation a character appears that has a beneficial influence on the individual, the chance that the individual will survive is increased, not only for itself, but for all of its descendants that come to inherit this character. It is this increase in the number of individuals possessing a particular character, that might have an influence on the course of evolution.” (187-189)
This is an abbreviated framework for understanding evolution under the “new mutations” or “mutation-limited” view that is now commonplace in molecular evolution. A new mutation arises and may “become established- we would say “become fixed” or “reach fixation” in population-genetics jargon- with a probability (not a certainty) that depends on its effects. If its effects are injurious, is has practically no chance of being established, and so on.
Morgan’s verbal description is remarkably accurate. Later, in the 1920s, Haldane, Wright, and Fisher began to work out some approximations for the probability of fixation of a new mutant allele. For newly introduced neutral alleles, P = 1/N (substitute 2N for diploids), where N is the population size, and this value is not affected by recessivity or dominance, just as Morgan says; for a newly introduced beneficial allele, P ~ 2s, where s is the selective advantage; for a significantly deleterious allele, the probability of fixation is vanishingly small. Later, diffusion theory was used to derive a more general expression for the probability of fixation (e.g., Gillespie, 1998, p. 82)
![]()
where the starting frequency p would be 1/N for a new mutation in the haploid case (and 1/2N for the diploid case).
To the extent that there was a distinctive “mutationist” perspective on evolutionary genetics that was rejected for its non-Darwinian implications, this was it. While Haldane, Fisher and Wright worked out the theoryA for the probability of fixation of a new mutation, they didn’t use this knowledge for anything important, because evolution by new mutations was not part of their theoryC of evolution. Instead, Morgan’s view of evolution as a series of mutation-fixation events was rejected by the Modern Synthesis as the “lucky mutant” view, and was ignored for nearly 50 years; Kimura popularized a neutral version of this view, which remained associated with neutral evolution for another 30 years; and in the past 10 years, Morgan’s perspective is emerging as a more general view that may serve as the basis for models of adaptation (e.g., Orr, 2002).
The Mendelian interpretation of continuous variation
The advocates of Darwin’s view of blending inheritance and fluctuation fought hard against Mendelism early in the 20th century, leading to the infamous biometrician-Mendelian debate. Thus, a century ago, it was necessary to defend Mendelian principles from attack by those who— disparaging Mendelism’s simplistic rules and suspecting its experimental foundations in “artificial” breeding— held out hopes for a fuzzier, more organic conception of heredity and variation that would fit better with Darwin’s view. In Bateson‘s 1902 “defense” of Mendelism, he provides a Mendelian interpretation of continuous variation:
“In the case of a population presenting continuous variation in regard to say, stature, it is easy to see how purity of the gametes in respect of any intensities of that character might not in ordinary circumstances be capable of detection. There are doubtless more than two pure gametic forms of this character, but there may quite conceivably be six or eight. When it is remembered that each heterozygous combination of any two may have its own appropriate stature, and that such a character is distinctly dependent on external conditions, the mere fact that the observed curves of stature give ‘chance distributions’ is not surprising and may still be compatible with purity of gametes in respect of certain pure types.” (p. 31)
By “chance distribution”, Bateson is invoking what we now call a “normal distribution”. Such a distribution “may still be compatible with the purity of the gametes”, i.e., compatible with Mendelian inheritance, because it can result by the combined effects of a multiplicity of Mendelian loci (6 or 8, he imagines), each with 2 homozygotes and 1 heterozygote, with environmental variation due to “external conditions”.
Thus, Bateson interpreted quantitative characters precisely as we do today, as the result of overlaying environmental fluctuation on a discrete distribution of genetic types. This interpretation is not due to little Ronny Fisher, the 12-year-old boy who would grow up to be a founder of mathematical population genetics and would declare that genetics was the key to Darwin’s theory7, but to Bateson and other geneticists, including Danish botanist Wilhelm Johannsen and the Swedish geneticist Herman Nilsson-Ehle.
The Mendelian interpretation was bolstered by a series of precise quantitative experiments conducted by Johannsen with the Princess bean. Johannsen isolated 19 stable self-fertilizing lines, each of which produced seeds with a different average weight. Planting any single variety would produce a smooth distribution of seed weights. Johannsen selected larger beans to plant a new generation, but this had no significant effect on the distribution of seed weights, proving that this newly arising variation was not heritable Darwinian fluctuation, but non-heritable somatic variation. Johannsen coined the terms “genotype” and “phenotype” to help explain this distinction.
By 1909, both Johannsen and Nilsson-Ehle had contrived to generate populations that, at the level of “genotype”, were known mixtures of discrete Mendelian types, but which- at the level of “phenotype”- produced a nice smooth bell-shaped distribution. Johannsen’s distributions of beans are reproduced in the figure below (right) from Morgan (1916; online source). 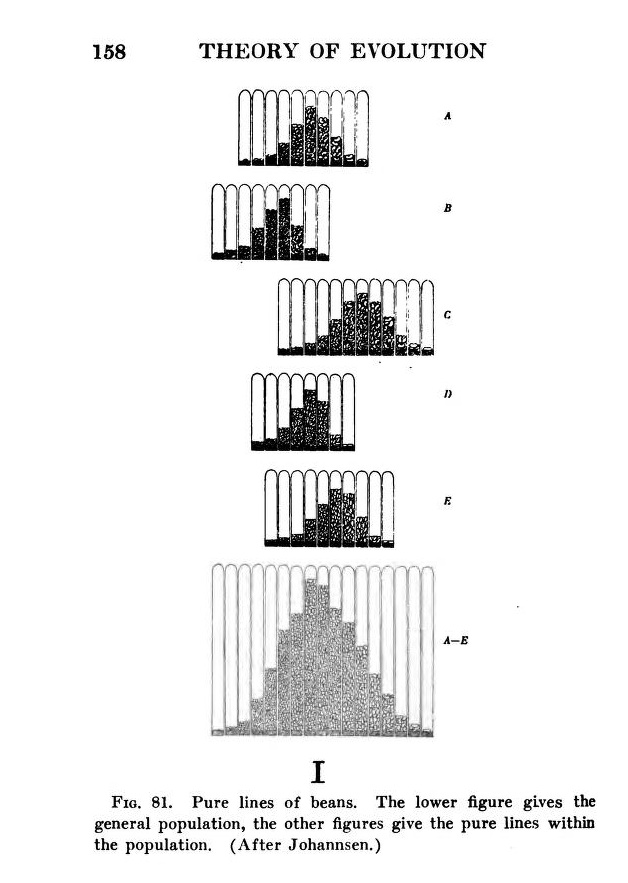
The evolution of quantitative characters
Finally, the Mendelians developed a causal theory for the gradual change in a quantitative character due to selection that negotiated the phenotype-genotype distinction and was appropriately probabilistic.
In the Darwinian view based on fluctuation and blending of hereditary substances, the superficial appearance that the whole population has shifted continuously and homogeneously reflects the underlying reality that hereditary substances have shifted continuously and homogeneously.
The new Mendelian view differed in two respects. First, given the genotype-phenotype distinction, selection of a particular phenotypic range implicates hereditary factors indirectly and probabilistically. For instance, Punnett (1911) constructs a simplified example in which there are just 3 genetically defined types, A, B and C, with mean weights of 10, 12 and 14 grains (a “grain” is a unit of weight equal to 0.065 gram). “A seed that weighs 12 grains may belong to any of these three strains. It may be an average seed of B, or a rather large seed of A, or a rather small seed of C” (p. 162; online source):
“On this view we can understand why selection of the largest seed[s] raises the average weight in the next generation. We are picking out more of C and less of A and B, and as this process is repeated the proportion of C gradually increases and we get the appearance of selection acting on a continuously varying homogenous material and producing a permanent effect.”
Second, as the Mendelians stressed repeatedly, the end result of this process is a not a new complement of hereditary factors, but a mixture of old components in new and different proportions. The hereditary factors are not changed by this process (as Darwin and his followers wrongly believed): only their proportions in the population are changed. Without new mutations, the new population would never transcend the genetic limits inherent in the original mix.
<!–
Homework
The popular view of history reflected in the Mutationism Myth is that our contemporary understanding of evolution began with Fisher, Haldane and Wright, not with Bateson, Morgan, Johannsen, Punnett and others. I see this as a whitewashed version of history, in which the contributions of the Mendelians have been erased.
But lets consider for a moment that, just as Darwin’s followers did not give up on blending inheritance without a nasty fight that created lasting suspicions about geneticists, they are not likely to give up Synthesis Historiography8 without a nasty fight that will leave a stain on critics such as myself. So, how does one convincingly establish a point about influence or credit? How do we know whose views were influential and whose views were purged? Here are some examples of types of information that might be useful:
- A popular evolution education web site has a timeline listing important contributors to evolutionary thinking. The timeline has a gap of a whole generation between the late-19th-century neo-Darwinians (e.g., Weismann) and the early “Synthesis” architects. Kimura is not listed.
- Morgan published several books on evolution that went through multiple printings; the Boston Public Library includes his 1916 book in its list of 100 most influential books of the 20th century.
- The Oxford Encyclopedia of Evolution, which includes biographic entries, does not have an entry for any Mendelian except Morgan, whose evolutionary views are not discussed.
In what other ways might we establish objectively that certain scientists, and not others, receive credit for their work and get included in histories? How could you generate a large amount of data quickly? How could one show an unwarranted or extra-scientific bias for or against certain authors, i.e., excluding the alternative possibility that “the judgment of history” favoring one person over another reflects true scientific merit?
–>
Conclusion
The Mutationism Myth suggests that our contemporary understanding of evolution did not emerge until Fisher, Haldane and Wright combined Darwin’s principle of selection with Mendelian genetics; and that a generation was wasted while Mendelians developed a “doomed rival” to Darwin’s great theory, in the form of a “mutationist” view that denied selection.
In fact, the Mendelians did not develop such a view. Instead, their interpretations paved the way for the Modern Synthesis and laid the foundations for our contemporary genetics-based understanding of evolution: they developed the Hardy-Weinberg model, interpreted quantitative trait evolution correctly, and even thought ahead to the “new mutations” perspective currently making inroads into evolutionary genetics.
Among the Mendelians, I also would count Nikolai Vavilov, the extraordinary Russian geneticist who started the first global seed bank (which persists today at the Vavilov Institute), leading expeditions that collected some 200,000 seeds. In 1922 he made a fascinating contribution to “mutationist” thinking, proposing parallel variations as a key component of parallel evolution. Vavilov was sent by the Soviets to a prison camp, where he died in 1943.
The Soviets purged Vavilov because of his opposition to Lysenkoism, the non-Mendelian theory of genetics with a Lamarckian theme of improvement-through-effort that fit nicely with Soviet ideology. Why were the contributions of Mendelians purged from our history, leaving the false impression of a generation-long gap in our intellectual history? Why don’t we count Bateson, Morgan, Punnett, Johannsen, and others among the “founders” of modern evolutionary thinking? Possible answers to this question will emerge in part 4 of The Mutationism Myth, where we explore the non-Darwinian aspects of Mendelian thinking, and in part 5, where we consider the “Modern Synthesis” as a restoration of Darwinian orthodoxy.
References
Bateson, W., and E. R. Saunders. 1902. Experimental Studies in the Physiology of Heredity. Reports to the Evolution Committee. Royal Society. (Bateson%20saunders&pg=PP1#v=onepage&q&f=false”>online source)
Bateson, W. 1902. Mendel’s Principles of Heredity: A Defense. Cambridge University Press, Cambridge. (online source)
Bateson, W. 1909. Heredity and Variation in Modern Light. Pp. 85-101 in A. C. Seward, ed. Darwin and Modern Science: Essays in Commemoration of the Centenary of the Birgh of Charles Darwin and of the Fiftieth Anniversary of the publication of the Origin of Species. Cambridge, London.
Gillespie, J. H. 1998. Population Genetics: A Concise Guide. Johns Hopkins University Press, Baltimore, MD.
Morgan, T. H. 1932. The Scientific Basis of Evolution. W.W. Norton & Co., New York.
Orr, H. A. 2002. The population genetics of adaptation: the adaptation of DNA sequences. Evolution Int J Org Evolution 56:1317-1330.
Punnett, R. C. 1911. Mendelism. MacMillan. http://www.archive.org/stream/mendelism00punn#page/172
Stephens, C. 2001. Selection, Drift, and the “Forces” of Evolution. Philosophy of Science 71:550-570.
Sturtevant, A. H. 1965. The Early Mendelians. Proceedings of the American Philosophical Society 109:199-208.
Vavilov, N. I. 1922. The Law of Homologous Series in Variation. J. Heredity 12:47-89.
Notes
1 As quoted in part 1, mutationism is a source of “mirth” for Dawkins.
2 The words “doomed rivals” are also from Dawkins. Back when I was a lad in school, my evolution professor- Dr. Kenneth Christiansen, who has been at Grinnell College for at least 45 years and is still there today- had a slightly gentler way of referring to alternative theories as the “also-rans”.
3 Note that Morgan‘s choice of words leaves some wiggle room that some other kind of variability could be offered “in support of” Darwin’s theory. A century ago, admiration for Darwin was nearly universal, as it is today. The meaning of Darwin’s theory, and ownership of the “Darwin” brand, were contested by scientists. By 1950, the “Modern Synthesis” school had captured the Darwin brand and began to use it more aggressively than they had dared to do before. However, things might have turned out differently. De Vries labeled himself as a “Darwinian”. Bateson and others sometimes cozied up to “Darwinism”.
4 Sturtevant (1965) lists 22 Mendelians who published from 1900 to 1905, and notes that all but 5 were under 40 (the older exceptions are de Vries, Garrod, Johannson, Wilson and Lang).
5 “stability analysis” means finding the “attractors” or points of stability in a dynamic system.
6 Hardy reveled in the purity of mathematics and stated that he had no desire to do anything useful. He said that his most important discovery was Srinivasan Ramanujan, a largely self-taught Indian genius who had written to Hardy and others seeking a mentor- only Hardy recognized his genius.
7 Synthesis Historiography attributes the resolution of Darwinism and Mendelism to Fisher (1918). In reality, the problem that Fisher (1918) solved was how to derive correlations of relatives as a formal consequence of Mendelian principles. Yule had proposed an analogous (flawed) resolution much earlier, in 1901, and it was immediately accepted by Bateson. Johannsen, Punnett and others clearly accepted selection on a range of small-effect variation. To draw attention to Fisher, suggests that, as late as 1918, “Darwinists” were still rejecting Mendelism in favor of blending, and needed to be told by one of their own that there was no contradiction. Interestingly, the biological argument that Fisher makes in his 1918 paper is mistaken. Roughly speaking, variation in quantitative traits is about half hereditary and half environmental, and there was evidence of this from before 1910, but Fisher argues in his 1918 paper that quantitative variation is nearly all hereditary, and the environmental component is negligible. This position was favorable to Fisher’s other views, of course.
8 “Synthesis Historiography” is Ron Amundson’s term for the industry of writing versions of history in which the Modern Synthesis is presented as the manifest destiny of science, and Mayr, et al are the heroes, while their intellectual opponents are fools and knaves.
*The Curious Disconnect is the blog of evolutionary biologist Arlin Stoltzfus, available at www.molevol.org/cdblog. An updated version of the post below will be maintained at www.molevol.org/cdblog/mutationism_myth3 (Arlin Stoltzfus, ©2010)