Unfamiliar ideas are often mis-identified and mis-characterized. It takes time for a new idea to be sufficiently familiar that it can be debated meaningfully. We look forward to those more meaningful debates. Until then, fending off bad takes is the order of the day! See the Bad Takes Index.
In his equally entertaining and obnoxious piece “The frailty of adaptive hypotheses for the origins of organismal complexity,” Lynch (2007) writes
The notion that mutation pressure can be a driving force in evolution is not new (6, 24–31)
citing works of Darwin, Morgan, Dover, Nei, Cavalier-Smith, and Stoltzfus and Yampolsky.
What does it mean to invoke evolutionary change due to a driving force of mutation pressure? This language suggests a process of population transformation by the mutational conversion of individuals, in contrast to population transformation by reproductive replacement.
That is, in a simplified world of discretely inherited types, we can imagine two general ways to transform a reproducing population from mainly type A to mainly type B. One mode is for an initially rare type B to take over the A population, over many generations, by the cumulative effects of differential reproduction, either biased (selection) or unbiased (drift). Individuals of type B over-produce, while A individuals die out, so that A individuals are replaced by unrelated B individuals. This is usually how we think about the transformation of populations: reproductive replacement. Selection and drift are often listed as the two main causes of evolution, and they act by reproductive replacement.
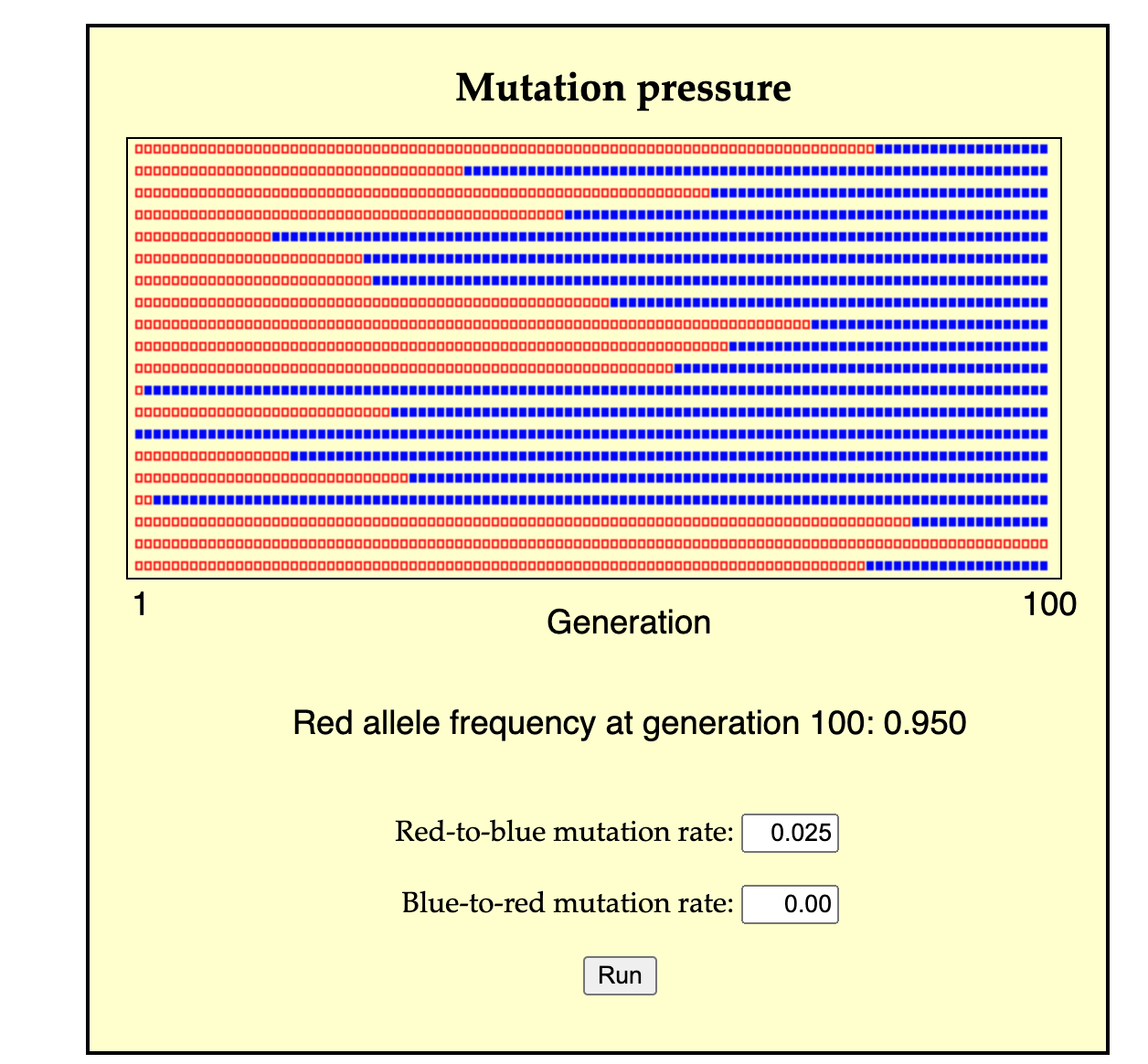
A second possible mode of change is for a population of predominantly A individuals to change by many separate events of A-to-B conversion (either individual conversion, or the cross-generational conversion of a lineage from parent to offspring). In this case, A individuals are lost, not by death, but by conversion, and likewise, B individuals are over-produced, not by the excess reproduction of B parents, but by conversion from A individuals. This process might take a single generation or many generations, depending on the rate of conversion (see image for a simulation).
A simulation of evolution by mutation pressure (from the mutation pressure page developed by John McDonald). N = 20 red squares at left represent individuals, with offspring generations going to the right. The reproductive variance is 0, so each individual leaves exactly 1 offspring in the next generation, inheriting the parental state or a mutated state with u = 0.025. By 100 generations, the population is mostly transformed, by mutational conversion alone, without reproductive differences.
In a more complex scenario, there are other possibilities. For instance, given diploid inheritance we could consider a process of biased gene conversion by which A1A2 genotypes are converted into A2A2. Suppose that A2 is recessive so that A1A1 and A1A2 have phenotype P1, and A2A2 has P2. In this scenario, biased gene conversion can transform a predominantly P1 population into a P2 population. Dover’s ideas about molecular drive combine effects of conversion and replacement.
One of the minor theories in Darwin’s Origin of Species is the mass transformation of individuals by direct effects of the environment. This idea was not unique to Darwin, but simply reflected 19th-century thinking by which heredity is (in effect) mediated by responsive memory-fluids that circulate in the body: after collecting bodily experiences, the memory-fluids gather in the gametes, and during reproduction, they blend, passing on a blended version of inheritance plus experience. Given this view, it was natural to suppose that, when animals or plants encounter a new environment, this results in a hereditary transformation by the cumulative effect of many environment-induced conversions.
The literature of the pre-Synthesis period includes some (typically ambiguous) references to population transformation by mutational conversion, e.g., Shull (1936) writes
If a given mutation were to happen often enough, and nothing opposed its survival, it could easily spread through the entire species, replacing all the other genes at the same locus.
Evolution by mutation pressure according to Haldane and Fisher
In the broader context of evolutionary theorizing, the mutation pressure theory appears most prominently as a strawman rejected by Haldane (1927, 1932) and Fisher (1930). That is, Haldane and Fisher did not advocate the notion of evolution by mutation pressure, but presented an unworkable theory as a way to reject the idea, popular among critics of neo-Darwinism, that evolutionary tendencies may reflect internal variational tendencies. In reality, the early geneticists typically argued, not for mutational transformation of populations, but for a two-step process of “mutation proposes, selection disposes” (decides); and the idea of orthogenesis was typically an appeal to what we might call “constraints” today. That is, the mutation pressure theory began as a dubious take on internalist thinking.
Regardless, Haldane and Fisher worked out the implications of evolution by mutation pressure, finding it unlikely on the grounds that, because mutation rates are small, mutation is a weak pressure on allele frequencies, easily overcome by opposing selection. Haldane concluded that this pressure would not be important except in the case of neutral characters or abnormally high mutation rates.
The conclusion of Haldane (1927)
To understand what Haldane is doing, one must bear in mind that, in the neo-Darwinian tradition, selection is the model of an evolutionary cause: other factors or processes are considered to be causal only to the extent that they look like selection. What selection does is to shift frequencies (and ultimately drive alleles to fixation), so Darwin’s modern followers define evolution as shifting frequencies and they define causal forces as pressures that might cause fixation. In effect, Haldane equates the importance of mutation with the potential for mutation pressure to drive allele frequencies. In this way of thinking, if mutation-biased evolution is happening, this is because mutation is driving alleles to high frequency against the opposing pressure of selection, which leads to Haldane’s conclusion that either (1) the mutation rate has to be abnormally high, or (2) selection has to be practically absent (i.e., neutrality). Fisher’s (1930) reasoning was similar. From the observed smallness of mutation rates, he drew a sweeping conclusion to the effect that internalist theories are incompatible with population genetics.
Provine (1978) identifies this argument (against evolution by mutation pressure) as one of the key contributions of theoretical population genetics to the Modern Synthesis, because it gave Darwin’s followers a seemingly rigorous basis to reject internalist theories (establishing the core Synthesis principle of There Is No Alternative). The argument was cited repeatedly by the architects of the Modern Synthesis (for examples, see Stoltzfus, 2017), and continues to be cited, e.g., Gould (2002) cites Fisher’s version of the argument and concludes that
Since orthogenesis can only operate when mutation pressure becomes high enough to act as an agent of evolutionary change, empirical data on low mutation rates sound the death-knell of internalism. (p. 510)
Subsequent work has partially undermined the narrow implications of the Haldane-Fisher argument, and completely undermined its broader application as a cudgel against internalism. Mutation pressure is almost never a reasonable cause of population transformation, because it would happen so slowly and take so long that other factors such as drift would intervene, as argued by Kimura (1980). The case studied by Masel and Maughan (2007) is a rare example in which evolution by mutation pressure is reasonable: the authors estimate an aggregate mutation rate of 0.003 for loss of a trait (sporulation) dependent on many loci, concluding that complex traits can be lost in a reasonable period of time due primarily to mutational degradation.
Thus, in spite of what one would conclude from Haldane (1927), patterns of mutation bias in evolution generally do not indicate evolution by mutation pressure via high mutation rates, or via neutral characters. Mutation-biased neutral evolution happens, not because mutation pressure is driving alleles to fixation in a biased way (instead, drift is the cause of fixation), but due to a bias in the origination process. And of course, Yampolsky and Stoltzfus (2001) showed that, when there is a bias in the introduction process, this can impose a bias on the course of evolutionary change even when fixations are selective, i.e., there is no requirement for neutral evolution.
In summary, the classic theory of evolution by mutation pressure is not much use in understanding evolution, and is mainly of historical interest for its role in an influential fallacy: generations of evolutionary thinkers believed wrongly that the mutation pressure theory proves mathematically that internalist theories are incompatible with population genetics.
Other theories
Now, with this background, we may return to Lynch’s bad take, associating various authors with mutation pressure as a driving force. Of the authors cited — Darwin, Morgan, Dover, Nei, Cavalier-Smith and my colleagues and I — none of them directly propose a theory of evolution by mutation pressure. However, the ideas of Darwin and Dover depict a process reliant on mass conversion: in Dover’s case, population transformation takes place by a dual process of conversion (gene conversion or sub-genomic replication) and reproductive replacement, and in Darwin’s case, it takes place by direct inherited effects of the environment.
Nei refers to “mutation-driven” evolution (the title of his 2013 book), but this is not a reference to mutation driving alleles to fixation. Nei’s usage of “drive” is descriptive or explanatory: evolution is mutation-driven to the extent that our understanding of important aspects of the course of evolution relies on knowing which mutations happen at what times. The same meaning is used in “Mutation-Driven Parallel Evolution During Viral Adaptation” (Sackman, et al. 2017). For an explanation of this meaning of “drive,” see Bad Take #4.
Likewise, the work from my colleagues and me is not about evolution by mutation pressure. From the very beginning, we have (1) followed Provine (1978) in noting the historical importance of the Haldane-Fisher argument against evolution by mutation pressure, and (2) promoted a theory for the effects of biases in the introduction process, obviously a different theory because it contradicts the implications of the mutation pressure theory.
So, what on earth does Lynch mean when he refers to evolution driven by mutation pressure? This is unclear. The model that Lynch (2007) presents immediately after the quoted statement is not a model of evolution by mutation pressure in the classic sense of Haldane and Fisher and IMHO does not correspond to what any of the cited authors are trying to say.
To understand what the model tells us, we must analyze it in detail, in comparison to the classic mutation-selection balance (also co-developed by Haldane and Fisher). The forces of population genetics are conceptualized like the laws of statistical physics, as mass-action pressures on allele frequencies due to the aggregate effect of countless individual events. In the case of mutation, countless individual events of mutational conversion from allele A1 to allele A2 result in a force or pressure of mutation shifting quantities of A1 to A2. Because there are innumerable independent events, each with an infinitesimal effect, we can represent the aggregate effect with a continuous quantity, e.g., we can write fA2‘ = fA2 + u fA1 to indicate the increase in fA2 due to mutation at rate u from allele A1, and we can write fA1‘ = (1 – u) fA1 to represent the corresponding reduction in the frequency of allele A1 due to mutation to allele A2.
In the classic conception of the mutation-selection balance, if A1 is favored over A2 by a selection coefficient s, then reproductive replacement by selection represents a pressure of magnitude s increasing fA1 and decreasing fA2, whereas mutation is a pressure of magnitude u with the opposite effect, acting by conversion (rather than reproductive replacement). The equilibrium frequency of A2 is roughly f = u / s, and this is typically a small number (much closer to 0 than to 1) because mutation rates are very small, e.g., a typical rate for a specific nucleotide mutation is 10-9 per generation. This is why Haldane concluded (above) that mutation would be unimportant unless selection is effectively absent (i.e., neutrality) or mutation rates are abnormally large (note how the classical mutation-selection balance of Haldane and Fisher is closely related to their argument about evolution by mutation pressure).
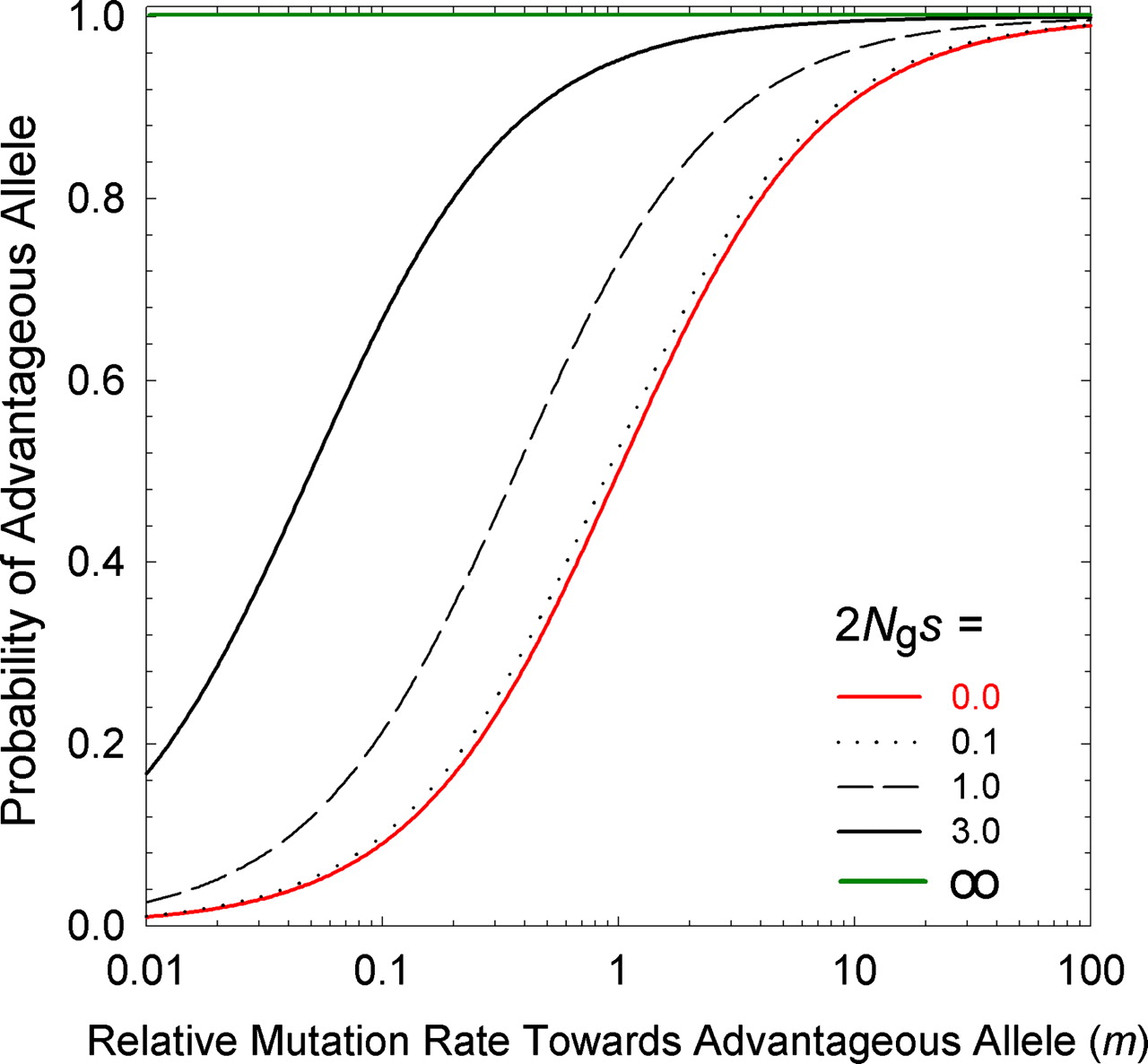
Lynch appears to reach a different solution to the same problem of the equilibrium frequency in a 2-allele system. His equation for the ratio of A1 to A2 is meS, where m is the forward-backward mutation bias favoring A1, and eS is the ratio of fixation probabilities where upper-case S = 2Ngs and lower-case s has the same meaning as above. However, this result actually does not represent the equilibrium frequency of A1 in a population of individuals, as for Haldane-Fisher: instead, it refers to the equilibrium distribution of infinitely many loci subject to an origin-fixation process, where each locus is fixed for A1 or A2, that is, meS is the expected ratio of (1) the fraction of loci fixed for A1 to (2) the fraction of loci fixed for A2.
(Figure 1 of Lynch, 2007)
This is easier to understand with a concrete example: the relative genomic frequency of two synonymous codons like CAT and CAC encoding histidine, where one of the codons (let’s assume CAC) is slightly more favored by selection.
The two cases, classic and Lynch, correspond to the large-population and small-population approximations for the ratio of favored to disfavored codons in Bulmer’s (1991) mutation-selection-drift model. In a large deterministic population, each histidine codon is fixed for the favored synonym (CAC), yet the disfavored codon (CAT) is maintained at low frequency in mutation-selection balance per Haldane-Fisher. In the small population, each histidine codon is fixed for either the favored codon (CAC) or the disfavored codon (CAT), and the frequency distribution of fixed states for loci is determined by the balance of two origin-fixation rates. So, in either case, if the disfavored codon is expected a fraction f of the time, then f is also the expected frequency of that codon over an infinite genomic set of histidine sites.
Thus, Lynch’s argument gives a different result because it refers to a different kind of mass-action pressure than Haldane and Fisher conceived. The relevant pressure in Lynch’s argument is the mass-action pressure due to events of origination aggregated over an infinite distribution of loci (sites). This origination pressure is not the same as classic mutation pressure, which is the mass-action pressure due to mutational conversion events aggregated over infinitely many alleles (in a population) at the same locus.
The result of this pressure (relative to a deterministic universe with only the favored codon), is to ensure that, for small values of S = 2Ngs, a substantial fraction of loci are fixed for the disfavored state; when S = 2Ngs becomes modestly large, this fraction is negligible. That is, mutation pressure, for Lynch, refers to something that ensures the predictable presence of deleterious states. By contrast, the theory of Yampolsky and Stoltzfus (2001) is about the way that biases in origination impose biases on which path, out of many possible, is taken by adaptation.
Clearly this model ensures the presence of deleterious states for small values of S, but it is not clear what justifies Lynch’s framing of this as an effect of a pressure of mutation (more precisely, a pressure of origination), rather than as an effect of random drift or of origin-fixation pressure. Mutation and fixation do not act separately in the context of the argument, and drift is profoundly important in ensuring that, in Lynch’s stochastic anti-paradise, a substantial fraction of everything is in a sub-optimal state. In a world that has deterministic selection, the favored codon always wins (and the disfavored one is never fixed by chance), even in small populations, and this will be true regardless of what we assume about mutation. Metaphorically, mutation is just knocking at the door, offering bad choices: drift has to open the door and let them in. On this basis, Lynch ought to point the finger at drift (not mutation) as the reason for non-optimality.
In fact, this is all utterly misleading when taken in context. After telling the reader that the idea of evolution by mutation pressure is not new, Lynch continues as follows
The notion that mutation pressure can be a driving force in evolution is not new (6, 24–31), and the conditions that must be fulfilled if mutation is to alter the direction of evolution relative to adaptive expectations are readily derived.
This is a sweeping claim about mutation and directionality! Yet, what follows is not a general model of effects of mutation on the direction of evolutionary change, but Bulmer’s model featuring fixations of deleterious alleles! That is, Lynch refers generally to mutational effects altering the “direction” of evolution, yet apparently, given his Manichean worldview, “direction” is just a matter of down vs up in fitness. And we just established that drift, not mutation, is the cause of deleterious fixations, which will happen even if there is no mutation bias (e.g., the disfavored codon will sometimes be fixed, even without any mutation bias). In the end, Lynch has presented correct mathematical results, but framed these results in an incorrect way that can only lead to confusion.
Accordingly, Svensson (here or here) has repeatedly claimed, citing Lynch’s paper, that an effect of mutation bias on adaptation would require “drift in small populations.” This error arises from a literal reading of Lynch, who (1) conflates diverse ideas (including ours) under the heading of evolution by mutation pressure, and then (2) makes a sweeping reference to mutational effects on “direction.” However, as explained, the effect requiring small populations in Lynch’s model is the fixation of a slightly deleterious allele by drift in small populations, whereas arguments about mutation-biased adaptation do not involve fixations of deleterious alleles at all, e.g., the behavior of the Yampolsky-Stoltzfus model does not rely in any sense on the fixation of deleterious alleles by drift.
What is the cause of so much misapprehension? The molecular revolution induced profound changes in thinking that have not been properly processed. Instead, we have attempted to squeeze a new understanding into the same old vocabulary— using old words for new concepts. In some cases, the result is verbal violence, as in the way that “Darwinian adaptation” is now used for the lucky mutant view previously known as a non-Darwinian theory of pre-adaptation. Familiar words are now overloaded with different concepts, and we have not paid attention to the problems caused by this overloading. Evolution by mutation pressure, in the classic Haldane-Fisher sense, means something different than what Lynch’s model means, which is something different than what the Yampolsky-Stoltzfus model means. The forces theory is inadequate, and leads scientists to incorrect conclusions, e.g., the assumption that mutation-biased evolution requires neutrality, which is pervasive in the literature.
The path toward greater clarity depends on making distinctions, e.g., distinguishing the introduction (origination) process from classical mutation pressure (across infinitely many copies of an allele in a population) and from origin pressure (across infinitely many loci). The reason to distinguish these, again, is that they behave differently, so that the rules for reasoning about one kind of causal process are different from the rules for reasoning about another.
Likewise, one must bear in mind that biological processes are not the same as the operators in models or mathematical formalisms, which capture only some of implications of biological processes for evolution. The classic conception of forces in population genetics includes a thing with the label “mutation” (and another thing with the label “selection”), but this thing does not have all the same implications as the biological process with the label “mutation.”
Sources that fail to make such distinctions will mislead readers with the impression that every reference to mutation is a reference to exactly the same evolutionary theory, when this clearly is not the case.
References
Fisher RA. 1930. The Genetical Theory of Natural Selection. London: Oxford University Press.
Gould SJ. 2002. The Structure of Evolutionary Theory. Cambridge, Massachusetts: Harvard University Press.
Haldane JBS. 1927. A mathematical theory of natural and artificial selection. V. Selection and mutation. Proc. Cam. Phil. Soc. 26:220-230.
Haldane JBS. 1932. The Causes of Evolution. New York: Longmans, Green and Co.
Haldane JBS. 1933. The part played by recurrent mutation in evolution. Am. Nat. 67:5-19.
Kimura M. 1980. Average time until fixation of a mutant allele in a finite population under continued mutation pressure: Studies by analytical, numerical, and pseudo-sampling methods. Proc Natl Acad Sci U S A 77:522-526.
Lynch M. 2007. The frailty of adaptive hypotheses for the origins of organismal complexity. Proc Natl Acad Sci U S A 104 Suppl 1:8597-8604.
Provine WB. 1978. The role of mathematical population geneticists in the evolutionary synthesis of the 1930s and 1940s. Stud Hist Biol. 2:167-192.
Shull AF. 1936. Evolution. New York: McGraw-Hill.
Stoltzfus A. 2006. Mutationism and the Dual Causation of Evolutionary Change. Evol Dev 8:304-317.
Yampolsky LY, Stoltzfus A. 2001. Bias in the introduction of variation as an orienting factor in evolution. Evol Dev 3:73-83.
Notes
Biased gene conversion is a newly recognized population-genetic force, non-identical with mutation, selection, or recombination. BGC is a conversion mechanism, not a replacement mechanism, but the formula for BGC is A1 + A2 –> A2 + A2, whereas the mutation formula is A1 –> A2. Similarly, the crossing-over formula is A1B1 + A2B2 –> A1B2 + A2B1. Thus, although the molecular operation of gene conversion is associated with cross-overs and with the machinery for recombination, the genetic operation of BGC is not the same thing as recombination.
Unfamiliar ideas are often mis-identified and mis-characterized. It takes time for a new idea to be sufficiently familiar that it can be debated meaningfully. We look forward to those more meaningful debates. Until then, fending off bad takes is the order of the day! See the Bad Takes Index.
In regard to reports of mutational biases influencing the changes involved in molecular adaptation, Svensson and Berger (2019) write
Despite the importance of mutations in these two studies, we emphasize that selection ultimately drove these adaptive allele frequency changes, rather than evolution being ‘mutation-driven’ as some might claim [1,7,8,13].
Actually the “mutation-driven” language is advocated in reference #1 (Nei’s book), but not in the other 3 sources cited, which are Yampolsky and Stoltzfus ( 2001), Stoltzfus (2006) and Stoltzfus and Cable (2014).
The authors object that, whereas the term “drive” refers to a cause that drives an allele to fixation, the changes implicated in the cited studies reflect selective fixation rather than fixation by mutation. The implication is that sources 1, 7, 8 and 13 advocate a theory of population transformation, not by reproductive replacement (via selection or drift), but by mutation pressure, i.e., the cumulative effect of many events of mutational conversion, which is generally a bad idea for reasons pointed out by Kimura (1980), although there are cases where it makes sense, e.g., loss of a complex character (for a more thorough explanation, see Bad take #2).
But of course, fixation by mutation pressure is not the theory advocated in Nei’s 2013 book Mutation-Driven Evolution, nor the other sources cited, nor sources such as this (note the title):
Nor is this what Pennings, et al. (2022) mean when they clarify that “our study is focused on the dynamics of adaptation and reversal in the context of point mutation-driven, stepwise evolution, rather than evolution through horizontal gene transfer or plasmid conjugation”. Nor is this what Tenaillon (2014) means when he writes
“In particular, the long-term evolution of 12 replicate populations of Escherichia coli by R.E. Lenski unraveled a succession of mutation fixations that reached up to 10 % fitness effect (Lenski & Travisano 1994). Large effect mutations appeared therefore to be the drivers of adaptation.”
Furthermore, Svensson and Berger (2019) surely know that authors such as ourselves or Masatoshi Nei, a famous population geneticist, are not advocating fixation by mutation pressure rather than by selection (or drift). For instance, the equation from Yampolsky and Stoltzfus that they recreate in Box 1 is based explicitly on the probability of fixation for a beneficial allele given by Haldane (1927).
That is, Svensson and Berger are making what is called a “bad-faith argument”, an argument that they know is wrong but which they use anyway, trusting that the argument gain favor with naive readers.
One must remember that the piece by Svensson and Berger (2019) is not a serious scholarly analysis, but a parody of Bad Synthesis Apologetics, a Sokal’s hoax exposing that— when the topic is either post-modernist cultural analysis or the status of evolutionary theory— it is possible to “publish an article liberally salted with nonsense if (a) it sounded good and (b) it flattered the editors’ ideological preconceptions.”
With their cheeky “mutation-driven” objection, the authors are parodying the kind of bad-faith argument that does not address any genuine issue of dispute, but is simply a way to score points with the kind of guileless reader who thinks Masatoshi Nei needs a lesson in basic population genetics from Svensson and Berger. It is a long-standing part of Synthesis culture to believe that critics of orthodoxy behave irrationally and hold views with obvious flaws.
If we take away this false pretense, the remaining issue is semantic: (1) does “mutation-driven” refer distinctively to the case in which mutation is a cause of allele fixation, i.e., the mutation pressure theory of evolution, or (2) does an additional meaning of “driving” exist that is more explanatory, justifying the use of “mutation-driven” for the case in which character and timing of evolutionary change depends on the character and the timing of mutations.
The issue is readily resolved by examining the usage of “drive” in evolutionary discourse. Does the literature of evolutionary biology restrict the “drive” language to causation only? The answer is clearly negative. Here is a tiny sample of recent uses from the technical literature:
Population size is clearly a condition, not a change-making causal process. Therefore, when our colleagues refer to population size “driving” something, this indicates an explanatory and not causal-mechanistic meaning of “drive.” The non-causal nature is unmistakable in the first example above, because what is being “driven” by population size is model choice, which does not physically exist in the realm of biology, but represents an abstraction in the realm of modeling. A cause X and its direct effect Y must occur in the same place, the locale of causation.
Note that this meaning of “drive” can be used — and often is used — with the concept of selection, i.e., we can talk about selection driving a thing, without that thing being an allele frequency, e.g.,
More generally, based on a purely descriptive analysis of patterns, e.g., a statistical analysis, scientists may refer to the predominant explanatory factor as the factor that “drives” the pattern. In this kind of claim, the implied chain of causation may be absent or unclear. As argued by Green and Jones (2016) in regard to “constraints,” scientists sometimes prefer a non-mechanistic language, because this allows them to discuss formal relations applicable to some system, without having to commit to a (potentially problematic) hypothesis for a mechanistic cause.
This does not mean that all uses of “drive” are equally welcome. When some authors above write that “These properties — and not function — seem to be the forces driving much of protein evolution” they are literally saying that properties are forces, which is gibberish. I find many of these uses of “drive” to be unhelpful, especially when results could be described more clearly using causal language (but see below).
To summarize, in their parody of Synthesis sophistry, Erik and David cover the “mutation-driven” issue with a delightfully empty misrepresentation sandwich, layered with bogus arguments. The meat is a weak semantic argument to the effect that the word “drive” must refer to population-genetic cause in the classic sense, a mass-action pressure that might cause allele fixation. Examples from the research literature demonstrate conclusively that the word “drive” simply does not have this restriction. This nutrient-poor semantic filling is sandwiched between two misrepresentations of the cited sources: (1) that they advocate the “mutation-driven” language (this is false for 3 of the 4 sources cited), and (2) that they invoke mutation pressure as a cause of fixation (this is false for all 4 sources).
Finally, note that we are having this discussion about language precisely because our customary causal language is insufficient. In the shifting-gene-frequencies theory of the Modern Synthesis, evolutionary causes are mass-action pressures (per statistical physics) that may cause allele fixations, e.g., selection and drift are seen as causes because they are potential causes of fixation. This theory of causes makes no distinction between shifts of a frequency from 0 to 1/N (or 1/(2N)) vs shifts among non-zero frequencies. When Haldane (1927) and Fisher (1930) addressed the potential for mutation-induced trends, they treated mutation as a cause of mass shifting and dismissed it as unimportant.
We have no other recognized causal language than statistical “forces” (“pressures”) at the population level. In particular, we have no recognized causal language for the effects of the introduction process: such effects are most often mis-described in terms of mutation pressure, or they are described indirectly or passively, as a matter of background conditions, or using the explanatory language of constraints or chance. The legacy of neo-Darwinism is that selection is the paradigm of a cause, and any other factor is judged to be causal or not depending on how much it acts like selection. Because the introduction process is not like selection at all, it has not been recognized as a causal process.
Attempts to describe the role of mutation actively rather than passively, with strong verbs, are certain to provoke opposition from the reactionary elements parodied by Svensson and Berger (2019). As I have written elsewhere, this position is cultural, not scientific: the reactionaries are culturally rigid but scientifically flexible. They will accept saltations (non-infinitesimal changes, major-effect alleles) and orthogenesis (tendencies due to internal biases) if the evidence demands it, but they will never endorse the terms “saltation,” “internal biases in evolution” or “orthogenesis,” because this would reveal a heretical departure from tradition. They will not reject mutation-biased adaptation due to biases in the introduction process, but they will describe it with old words while referencing dead authorities, in order to anchor new concepts in traditional sources (see also Bad Takes #5).
References
Kimura M. 1980. Average time until fixation of a mutant allele in a finite population under continued mutation pressure: Studies by analytical, numerical, and pseudo-sampling methods. Proc Natl Acad Sci U S A 77:522-526.
Unfamiliar ideas are often mis-identified and mis-characterized. It takes time for a new idea to be sufficiently familiar that it can be debated meaningfully. We look forward to those more meaningful debates. Until then, fending off bad takes is the order of the day! See the Bad Takes Index.
A common “stages of truth” meme holds that successful disruptive ideas are first (1) dismissed as absurd, then (2) resisted— the idea is declared unlikely and the evidence is strenuously disputed—, and finally (3) regarded as trivial and attributed to long tradition. Haldane’s version is that “The process of acceptance will pass through the usual four stages: (i) this is worthless nonsense; (ii) this is an interesting, but perverse, point of view; (iii) this is true, but quite unimportant; (iv) I always said so.” The QuoteInvestigator piece on the stages-of-truth meme has this version:
For it is ever so with any great truth. It must first be opposed, then ridiculed, after a while accepted, and then comes the time to prove that it is not new, and that the credit of it belongs to some one else
Svensson and Berger (2019)— in an article that reads like a Sokal’s hoax of Bad Synthesis Apologetics— model all the stages of truth in the same paper: (1) they dismiss strawman versions (e.g., mutation as an independent cause of adaptation) as absurd (see Bad Takes #3 and Bad Takes #4), (2) they present a clumsy version of the theory but dispute the evidence and declare it implausible based on a list of fake theoretical restrictions, and (3) finally, implicitly admitting that the phenomenon is real and that the theory we proposed is correct, they describe it as trivial and familiar:
These studies therefore only exemplify how historical contingency and mutational history interact with selection during adaptation to novel environments [31, 38, 52], entirely in line with standard evolutionary theory and the uncontroversial insight that different genomic regions contribute differentially to adaptation driven by selection, with mutations merely providing the genetic input [53].
In this way, the reader is guided through the stages of truth from patent absurdity to yesterday’s news.
However, our focus here is only on the end-point of this progression, in which Svensson and Berger (2019) give the impression that the new work on mutation-biased adaptation represents ordinary textbook knowledge, so that these new results induce no changes in evolutionary reasoning, raise no new questions, and suggest no new priorities for research. The specific implication of the passage above is that these findings are merely a matter of “contingency” and present nothing original or new relative to the contents of references 31, 38 and 52.
[figure legend: A recent exploration of “contingency” by Wong (2019), revealing the lack of a precise meaning other than something vaguely to do with chanciness.]
Yet contingency is not a causal theory: it is an explanatory concept indicating that a system is non-equilibrium, so that the state of the system cannot be predicted without knowing the initial conditions and detailed dynamics. The notion of contingency, by itself, does not provide a theory of the dynamics. If we try to answer the odd question, “what does contingency predict about how the mutation spectrum shapes the spectrum of adaptive substitutions?” then we will get nowhere without a theory for the dynamics, and this theory will have no need for a concept of contingency (an explanatory concept, not a cause of anything), but will directly addresses how the details of mutation rates influence the spectrum of adaptive substitutions.
Svensson and Berger have a bad habit of misrepresenting cited works. What do refs 31, 38 and 52 say? References 31 and 38 are from the field of quantitative genetics, and simply do not provide any such dynamical theory, e.g., here is the abstract to reference 31:
The introduction and rapid spread of Drosophila subobscura in the New World two decades ago provide an opportunity to determine the predictability and rate of evolution of a geographic cline. In ancestral Old World populations, wing length increases clinally with latitude. In North American populations, no wing length cline was detected one decade after the introduction. After two decades, however, a cline has evolved and largely converged on the ancestral cline. The rate of morphological evolution on a continental scale is very fast, relative even to rates measured within local populations. Nevertheless, different wing sections dominate the New versus Old World clines. Thus, the evolution of geographic variation in wing length has been predictable, but the means by which the cline is achieved is contingent.
Reference 52 is Good, et al (2017), a deep sequencing study of samples from Lenski’s LTEE (long-term evolution experiment). This is mainly an empirical analysis of allele trajectories and clonal interference and so on. There are no explicit claims for an effect of mutation bias on the spectrum of adaptive substitutions (mutation bias is mentioned only in relation to mutators, but mutators generate a lot of hitch-hikers in this experiment, so that the influence of mutators on the set of adaptive changes is not clearly established). Indeed, the presentation of results indicates in various places (e.g., the comments on parallelism) that Good et al are not paying attention to the issue of how mutation bias influences probabilities of beneficial changes.
What is going on here? Svensson and Berger (2019) seem intent on illustrating how to avoid addressing the novelty of (1) a formal pop-gen theory that focuses on the introduction process, and which makes novel predictions about evolution based on tendencies of variation (addressing aspects of parallelism, trends, GP maps, findability, etc), in a way that directly contradicts the classic Haldane-Fisher “mutation pressure” argument, and (2) empirical results confirming a distinctive prediction of this theory, namely effects of mutation biases on adaptation (not requiring neutrality or high mutation rates), contradicting a long neo-Darwinian tradition of dismissing internal biases in evolution.
One way to avoid these key issues is to engage in whataboutery, i.e., responding to an issue by demanding attention to a second issue. What about other research? What about selection? Whataboutery provides the writer an opportunity to engage the reader on some related topic, e.g., for purposes of name-dropping. Rather than taking the opportunity to educate readers on the details of a new and exciting — but poorly known — body of work on mutation bias and molecular adaptation, i.e., the ostensible topic of their commentary, Svensson and Berger instead lavish their attention on older and much better known work on related topics by eminent scientists, e.g., the LTEE from Lenski and colleagues, lizard stuff from Jonathan Losos, the famous stickleback Pitx1 example, or David Houle’s work on fly wings.
More generally, Svensson and Berger (2019) illustrate how Synthesis apologists do not contemplate the practice of science in terms of falsifiable theories, precise reasoning, or the prospect of striking future discoveries, but are mainly concerned with crafting a narrative of tradition that integrates important people and flexible themes. They trivialize new work by assigning it to familiar and vague categories that make it seem ordinary, rather than mapping it to the specific issues that motivate it, make it significant, and raise unanswered questions for the future.
Model of Bell’s first telephone from 1875
To understand how this game works, consider a completely unrelated example, namely the invention of a telephone 150 years ago (image). The novelty-hating curmudgeon may object as follows: You say there is something new here? How arrogant to make such a claim! There is nothing new here at all! This is merely an engineered device, and inventors have been crafting devices for centuries! I could show you 15 devices from just the past few years that are more impressive than this one, with more parts. You have done nothing to acknowledge this past work. Have you no respect? There is no fundamentally new technology here, merely pieces of wood and metal and wire! I could build something like this in an afternoon for $25. There are no new electrical or mechanical principles at work, merely electrical currents and vibrations controlled by magnets. It looks like other devices I have seen. I could break it easily with a hammer. I doubt that it can fly like an airplane.
The problem is not that these objections are false statements. They could all be true. The problem is that they fail to address the crucial issue: the telephone prototype instantiates a generalizable technology to support remote voice communication through wires, thus over long distances.
Svensson and Berger have done an excellent job of illustrating how to play the irrelevant-objections-to-novelty game. When they argue that new work on mutation-biased adaptation is just another example of contingency, this represents the strategy of describing new work in a trivially general way, like saying that the first telephone is just a device. When they claim that the theory we proposed is already part of the Modern Synthesis, on the grounds that it can be broken down into familiar parts, this is like objecting that the telephone is made of familiar parts and therefore does not represent something new but is merely part of a familiar tradition of constructing devices.
Of course, the significance of a new device— or a new theory— is not in the list of parts, but in what the assembled whole accomplishes.
What is the actual significance of recent work on mutation-biased adaptation? The essence of neo-Darwinism is a dichotomy of variation and selection, in which variation merely provides raw materials (substance, not form), and selection is the source of order, shape, and direction. Theories of evolution subject to internal biases directly contradict neo-Darwinism and were considered heretical. The argument of Haldane and Fisher that such theories are incompatible with population genetics (see Bad takes #2) was eagerly adopted by the architects of modern neo-Darwinism, yet (1) this classic conclusion is unwarranted theoretically and (2) its implications are refuted empirically. These two provocative claims are established by recent work on mutation-biased adaptation; they are not part of textbook knowledge; they are not established in well known studies cited by Svensson and Berger to illustrate scientific name-dropping.
Unfamiliar ideas are often mis-identified and mis-characterized. It takes time for a new idea to be sufficiently familiar that it can be debated meaningfully. We look forward to those more meaningful debates. Until then, fending off bad takes is the order of the day! See the Bad Takes Index.
A reviewer of Stoltzfus and Yampolsky (2009) wrote that “we have long known that mutation is important in evolution,” citing the following passage from Haldane (1932) as if to suggest that the message of our paper (emphasizing the dispositional role of mutation) was old news:
A selector of sufficient knowledge and power might perhaps obtain from the genes at present available in the human species a race combining an average intellect equal to that of Shakespeare with the stature of Carnera. But he could not produce a race of angels. For the moral character or for the wings, he would have to await or produce suitable mutations
We included this in the final version of the paper because, actually, this passage demonstrates the opposite of what the reviewer implies. What is Haldane suggesting?
I can’t resist a good story, so let’s begin with this 1930s photo of Italian boxer Primo Carnera, his friend and fellow heavyweight champ Max Baer, and Hollywood actress Myrna Loy. Baer dated Loy in real life. They made a movie together, the three of them (thus the staged publicity photo). Baer, one of the greatest punchers of all time and half-Jewish, became a hero to a generation of Jewish sports fans when he demolished Max Schmeling, the German champion, prompting Hitler to outlaw boxing with Jews. He literally killed one of his opponents, and repeatedly sent Carnera to the floor during their single fight.
Primo Carnera, Myrna Loy, and Max Baer in a publicity photo from the 1930s
But the point of this picture is that, although Baer was a formidable man, Carnera makes him look small. Other fighters were afraid to get in the ring with him. Though enormous — 30 cm taller and 50 kg heavy than the average Italian of his generation —, Carnera was not the aberrant product of a hormonal imbalance. This photo shows a huge man who is stocky but well proportioned, muscular, and surprisingly lean. Again, he was not a misshapen monster, but a man at the far extremes of a healthy human physique, which is precisely Haldane’s point.
Selective breeding to the quantitative extremes of known human ability, Haldane proposes, could produce a race combining the extreme of Carnera’s stature with Shakespeare’s magnificent verbal ability.
Haldane contrasts this with a different mode of evolution dependent on new mutations, which might produce a race of pure-hearted, winged angels, if one could wait long enough for the mutations to happen. That is, Haldane is contrasting (1) a mode of evolution that is gradual and combinatorial, bringing together known extremes, with (2) a mode of evolution that could generate imaginary fictitious not-real creatures. Haldane, Wright and Fisher each argued that a mode of change dependent on new mutations would be too slow to account for the observed facts of evolution. They argued instead that evolution must take place on the basis of abundant standing variation, a former orthodoxy that is largely forgotten today.
That is, in the passage above, Haldane is not endorsing a mode of mutation-dependent evolution, but gently mocking it, in contrast to a mode of evolution that, based on quantitative standing variation, could produce a race of magnificently eloquent champions.
Thus, the reviewer has missed Haldane’s meaning.
To understand what this reviewer is trying to accomplish via this “we have long known” argument, let’s imagine an alternative universe in which the reviewer says this:
“We have long known about the important role of biases in the introduction process emphasized in this manuscript. Haldane (1932) and Fisher (1930) explored the theoretical implications of such biases (under regimes of origin-fixation and clonal interference); Simpson and many others incorporated a theory of internal variational trends (i.e., orthogenesis) into their interpretations of the fossil record. Therefore, the authors’ implicit claim of novelty is unfounded. The theory is simply not new and not theirs, and they need to cite the proper sources for it.”
Of course, the reviewer does not say this, because nothing like this ever happened. In our universe, Fisher and Haldane failed to explore this theory (origin-fixation models didn’t appear until 1969, and clonal interference was not formally modeled until much later). In our universe, Simpson and others mocked the idea of orthogenesis.
Certainly, the reviewer is correct that scientists in the mainstream Modern Synthesis tradition have always known that mutation is important in evolution. Haldane, Fisher, Ford, Huxley, Dobzhansky, and others said explicitly that mutation is ultimately necessary, because without mutations, evolution would eventually grind to a halt.
However, they did not say that mutation is important as a dispositional factor. Instead, they argued explicitly against this idea, e.g., Haldane (1927) is the original source of the argument that mutation pressure is a weak force (see Bad takes #2).
The theory of biases in the introduction process, by contrast, says that mutation is important in evolution as a dispositional cause, a cause that makes some outcomes more likely than others, and that this importance is achieved (mechanistically) by way of biases in the introduction process.
So, the reviewer is doing a rhetorical feint (aka bait-and-switch argument): the words “we have long known…” encourage the reader to think that he is going to undermine the novelty of the theory, but his actual claim fails to do this. The theory of biases in the introduction of variation is a specific theory, linking certain kinds of inputs with certain kinds of outputs, via a certain kind of population-genetic mechanism. And the reviewer is responding to this theory by saying “we have long known that mutation is important” which is not the same thing. The words “mutation is important” do not by themselves specify this theory— or any theory—, and in fact, the traditional importance assigned to mutation is clearly not “dispositional cause that makes some outcomes more likely than others” but “ultimate source of raw materials without which evolution would grind to a halt.” These are two utterly different theories about the role of variation, and only one of them is traditional and neo-Darwinian.
Finally, it is important to understand the role of flimsy “we have long known” arguments in evolutionary discourse. The pattern of the argument is that it appears to undermine a claim of novelty by identifying the same claim in traditional sources, but what is actually happening is that a specific target is being swapped out for something else, often a fuzzy or generic claim. The novelty of X is rejected on the grounds that X sounds a lot like old theory Y, or because both X and Y can be categorized as a member of some larger and fuzzier class of claims, e.g., “chance” or “contingency” (see Bad Takes #5: Contingency). This is often the case with “we have long known” arguments emanating from traditionalist pundits.
Again, if a theory X is actually unoriginal, pundits don’t need to make vague “we have long known” arguments, but can simply cite the original source of X per standard scientific practice. It is precisely when X is new that traditionalist pundits must construct vague “we have long known” arguments to rescue tradition from its failures.
References
Haldane JBS. 1932. The Causes of Evolution. New York: Longmans, Green and Co.
Stoltzfus A, Yampolsky LY. 2009. Climbing mount probable: mutation as a cause of nonrandomness in evolution. J Hered 100:637-647.
Arlin Stoltzfus, 2012 to 2023 (@ArlinStoltzfus). Views expressed here are my own. [CC-BY ]