Why size matters: Saltationism, creativity, and the reign of the DiNOs
Debates on “gradualism” in evolutionary biology address the size distribution of evolutionary changes. The classical Darwinian position, better described as “infinitesimalism”, holds that evolutionary change is smooth in the sense of being composed of an abundance of infinitesimals (not one infinitesimal at a time, but a blending flow of infinitesimals). The alternative is that evolution sometimes involves “saltations” or jumps, i.e., distinctive and discrete steps. The dispute between these two positions has been a subject of acrimony at various times in the 20th century, with several minor skirmishes, and a larger battle with at least one genuine casualty (image).

Walter Frank Rafael Weldon (public domain image from wikipedia). Weldon ignored an advancing illness and worked himself to death (1906) poring over breeding records in an attempt to cast doubt on discrete inheritance. Along with Pearson and other “biometricians”, Weldon held to Darwin’s non-Mendelian view combining gradual hereditary fluctuations with blending inheritance.
Today, over a decade into the 21st century, we have abundant evidence for saltations, yet the term is virtually unknown, and we still seem to invoke selection under the assumption of gradualism. Are we saltationists, or not? I’m going to offer 3 answers below.
But first, we need to review why the issue is important for evolutionary theory.
Why size matters
Discussions of gradualism often seem obscure (who cares about size?) because they fail to explain that the distribution of sizes of variations is not the real issue, but merely the easiest way to relate crucial issues to something tangible and measurable. The crux of the matter is the extent to which an external force of selection, as opposed to an internal force of variation, can be understood as the cause of creativity, direction and initiative in evolution. The issues of how new things come about, what determines the direction of evolution, and what initiates change are weighty issues, worth fighting over. The answers to these questions matter: they not only affect our understanding of evolutionary causes, but they determine how we must conduct research if we wish to understand evolution properly.
How, exactly, does the size of variations relate to these big questions? If a distinctive new form is always connected to an ancestral form by a long series of infinitesimal shifts (gradualism), then the origin of the new form, and its distinctive character, are not explained by any one variation, nor even (to be precise) by a set of them, but by the process that accumulates so many individually trivial effects in one important direction.[1] If the variations fixed in evolution are individually tiny, and the guiding process is selection, then selection is creative, and we can say that selection is the cause of new traits because it composes them out of raw material. We can say, following Sterelny & Griffiths (1999), that “selection shapes organisms to their environment“. Mainstream evolutionary biologists remain committed to this view, which is why they reacted with such extraordinary hostility when Fodor and Palmarini-Piatelli tried to take away the idea of “selection for” as a valid form of explanation.
The role of variation in this view is necessary but trivial: if we wish to explain or predict the course of evolution, selection is what we must study, even though mutation is ultimately necessary. Studying mutation to understand evolution would be a fool’s errand, like studying gasoline to understand traffic patterns, or studying iron ore to understand modern architecture. This is the nature of a material cause: it provides substance only, not form or direction or initiative (the term “material cause” has a precise meaning in Aristotle’s four-fold taxonomy of causation).
That is, Darwinism is only secondarily a position on the sizes of changes fixed in evolution: it is primarily a position on the creativity and power of selection, and the corresponding lack of creativity and power of mutation. S. J. Gould was one person who understood this connection: he gives an extensive explanation on pp. 137 to 146 of The Structure of Evolutionary Theory. Darwin’s theory purports to replace a divine creator with a materialistic one. “Darwin’s theory therefore cannot be equated with the simple claim that natural selection operates”, but that “it acts as the creative force of evolutionary change” (italics original, p. 139). In order for selection to be creative in the way that Darwinism demands, variation must be (in Gould’s words) “copious”, “undirected”, and “small in effect.” At the risk of boring the reader, I’d like to quote from Gould at length (p. 140):
As the epitome of his own solution [to the thorny problem of creativity], Darwin admitted that his favored mechanism ‘made’ nothing, but held that natural selection must be deemed ‘creative’ (in any acceptable vernacular sense of the term) if its focal action of differential preservation and death could be construed as the primary cause for imparting direction to the process of evolutionary change. Darwin reasoned that natural selection can only play such a role if evolution obeys two crucial conditions: (1) if nothing about the provision of raw materials— that is, the sources of variation— imparts direction to evolutionary change; and (2) if change occurs by a long and insensible series of intermediary steps, each superintended by natural selection— so that “creativity” or “direction” can arise by the summation of increments.
Under these provisos, variation becomes raw material only– an isotropic sphere of potential about the modal form of a species. Natural selection, by superintending the differential preservation of a biassed region from this sphere in each generation, and by summing up (over countless repetitions) the tiny changes thus produced in each episode, can manufacture substantial, directional change. What else but natural selection could be called ‘creative,’ or direction-giving, in such a process? As long as variation only supplies raw material; as long as change accretes in an insensibly gradual manner; and as long as the reproductive advantages of certain individuals provide the statistical source of change; then natural selection must be construed as the directional cause of evolutionary modification.
These conditions are stringent; and they cannot be construed as vague, unconstraining, or too far in the distance to matter. In fact, I would argue that the single most brilliant (and daring) stroke in Darwin’s entire theory lay in his willingness to assert a set of precise and stringent requirements for variation— all in complete ignorance of the mechanics of heredity. Darwin understood that if any of these claims failed, natural selection could not be a creative force, and the theory of natural selection would collapse.
Note the top-down reasoning: Darwin decides what the role of variation must be in order for selection to be the powerful creative force that (in his view) it must be. This is especially poignant given that Darwin wrote 2 volumes on diverse kinds of variations, then set aside nearly all of that knowledge to build a theory just on “indefinite variability” a.k.a. “fluctuation”. This commitment to the subordinate role of variation is what makes Darwinism distinctive among theories of evolution. In all other theories, the process of generating variation has some dispositional role in influencing the outcome of evolution. In Darwinism, variation is merely a source of raw materials that selection shapes into adaptations.
What if evolution doesn’t work in this way? If evolutionary change comes, to some extent, in distinctive chunks (saltationism), each defined by a discrete event of mutation, we cannot talk about evolution in the same way, i.e., we can’t have the same kinds of causal explanations and hypotheses. To the extent that evolution depends on the introduction of rare variants (a key component of mutationism), we cannot treat the dynamics of evolution as being entirely externally driven. To the extent that change comes in chunks, things can’t be as exquisitely adapted as we might have believed otherwise.[1] To the extent that evolutionary change depends on discrete and distinctive variations, our causal explanation of the origin of new forms must reflect the internal process that generates distinctive variations. We can no longer use the “raw materials” metaphor and say that selection shapes organisms to their environments, as though it were working with malleable clay.
Finally, consider the implications for theories and research programs. If evolution takes place solely by infinitesimals, the sufficiency of evolutionary theory does not require any special theory of form and its variation— we only need to note that every trait varies infinitesimally, i.e., we just need something like the multivariate quantitative genetics of Lande & Arnold. But if not, then evolutionary theory is incomplete without a substantive theory of form and its variation. This realization, in turn, will shape how we do research: we are obliged to pursue the kind of research program that Bateson urged, in order to develop a substantive theory of variation and its propensities.
With that conceptual background, let us now consider whether the order of the day is gradualism, saltationism, or something else. I will give three different answers.
1. We’re saltationists
Forty years ago, Mayr and his cronies had the English-speaking world convinced that they had won the war against saltationism, and that the foolish views of de Vries, Bateson, Goldschmidt, and the early geneticists were dead. Leading researchers clearly no longer treat this as a dead issue. For instance, Stern (2000) explains the novel focus of evo-devo on “the problem of variation” by citing the view of de Vries, Goldschmidt and Bateson that “natural selection of small differences is insufficient to account for the large differences between taxa.” Orr (2005) complains that Fisher’s infinitesimal model stifled research for decades by “assuming away” the problems of adaptation, quoting Bateson’s sarcastic critique of this way of thinking:
By suggesting that the steps through which an adaptive mechanism arose were indefinite and insensible, all further trouble is spared. While it could be said that species arise by an insensible and imperceptible process of variation, there was clearly no use in tiring ourselves by trying to perceive that process. (Bateson 1909a)
Way back in 1992, gradualism was delivered a major blow by Orr and Coyne, whose abstract reads as follows:
We review both the theory and the data supporting the neo-Darwinian claim that adaptation results from the accumulation of many alleles of small effect. The theory supporting this “micromutationist” view is inconclusive. Furthermore, the evidence from genetic analysis is mixed: although some adaptations are apparently based on many genes of small effect, others clearly involve major genes. Most genetic analyses, moreover, have lacked the power to detect major genes. We conclude that, although the neo-Darwinian view of adaptation may be correct, it is not strongly supported by evidence. The introduction of molecular approaches may, however, finally allow us to resolve this question.
Reviewing evidence from the genetic dissection of inter-species differences— exactly the kind of evidence that neo-Darwinians confidently claimed in support of gradualism—, they argued that gradualism was unlikely, and that major effects were likely to be important. More interestingly, they concluded from historic evidence that the architects of the Modern Synthesis had not been justified to make the claims they had made originally. This 1992 review had a compromising tone (perhaps due to the dual authorship) that is absent in Orr’s more recent reviews, which describe the “micromutationist” (gradualist) position as “little more than a mathematical convenience” (Orr, 2005).
The case for saltationism in molecular evolution— easy to understand, and very persuasive— is argued in recent books by Koonin (2013) as well as Shapiro and Nei (see my reviews of the latter two here and here). We could argue this case from a variety of examples such as the origin of antifreeze genes in fish discussed by Logsdon and Doolittle (figure). There isn’t any room for quibbling about what represents a “slight” or “infinitesimal” (Darwin’s words) change when we get to sequences: any protein, RNA or genome clearly could have evolved by single-residue changes, e.g., ATGC can change to CAGTAC or TCAATGGA or any other sequence by a series of 1-residue changes. If evolution does not occur in this way, it is not gradual in the way Darwin and his followers clearly intended. In fact, genes often evolve via duplications; proteins often change by blocks being added or deleted; genomes undergo doublings and fusions. Molecular evolution often involves a jump, particularly in the incipient stages of useful structures— whereas the Darwinian position has always been that novelty does not require an initial jump (instead, a tiny shift in the right direction is enough to catch the attention of selection).
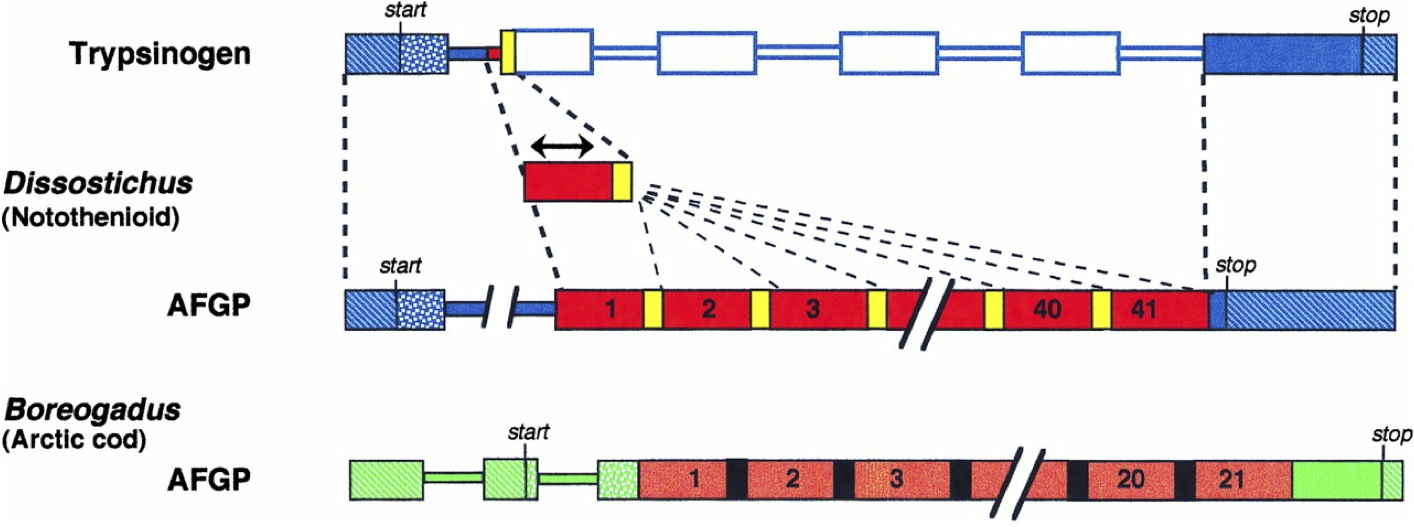
Discontinuous steps in the origin of antifreeze genes (figure from Logsdon and Doolittle, 1997).
As a result of such discoveries, scientists today have abandoned the Darwinian position on the underlying issues— how new things come about, and how to study evolution. The Modern Synthesis argued that, because variation is so abundant in the gene pool, it cancels itself out of the evolutionary equation. By contrast, one of the premises of contemporary evo-devo is that Bateson and de Vries were right: to understand innovation, we need to focus on the arrival of the fittest, not merely the survival of the fittest. Few evolutionary biologists support the classical position that natural selection is creative, which is no longer taught in textbooks. This doctrine is rejected explicitly in recent books about molecular evolution (e.g., Nei, Koonin) or evo-devo (e.g., Kirschner and Gerhardt).
Finally, we are starting to see the required shift in how we carry out research. Today, Bateson’s vision of a research program on new variation is being realized, e.g., in the evo-devo focus on candidate mutations, and in growing support for mutation-accumulation experiments (note that the study of new mutations is quite different from the Modern Synthesis focus on standing variation [2]).
2. We’re gradualists
No mainstream scientist today embraces the term “macromutation” or “saltation”. Whenever anyone invokes saltationism, macromutations or “hopeful monsters”, there is a backlash. In 2008 there was a piece by science journalist Olivia Judson that invoked “hopeful monsters”, and this provoked an apoplectic response from Jerry Coyne. When Nature published a newsy piece entitled “Evolution: Revenge of the hopeful monster“, Coyne was included to pose for gradualist orthodoxy.[3]
The reason for this is that the number of well documented exceptions to phenotypic gradualism uncovered by evo-devo and quantitative genetics in 25 years is small. When someone has to write a whole book to cover the topic, then it will be large enough to warrant some attention: today all the examples can be covered— and have been covered multiple times— in a single review paper. This is why, even in evo-devo, phenotypic variations are described as the raw materials for evolution.
The examples from molecular evolution could fill a book, but no one counts this as evidence against gradualism, because Darwinism only refers to big things we can see. Molecules are not important. For instance, to counter Koonin’s claim that endosymbiogenesis counts against gradualism, Forterre responds that the underlying molecular mechanisms are not what is important, but only the gross phenotypic changes, which probably were small. More generally, evolutionary discourse is bound by the Phenotypes of Large Animals rule, also known as the Darwin Couldn’t Have Known rule. Nothing is important in evolution unless it is visually obvious in classic study organisms. Nothing in molecular evolution can ever challenge neo-Darwinism.
More importantly, we remain committed to the Darwinian position on the underlying issues— how new things come about, and how to study evolution. When Kirschner & Gerhardt challenged the argument that selection is creative, Charlesworth rebutted their claims in a book review in Science, invoking the Modern Synthesis argument that selection creates improbable combinations out of the abundance of the gene pool. When Nei rejected this same argument as hand-waving, the book review by Wright (2013) in the conservative journal Evolution dismissed it, citing “persuasive arguments” made long ago:
Nei dismisses the notion that natural selection is a creative process as a rhetorical argument without grounding in either theory or empirical evidence, flying in the face of the persuasive arguments to the contrary advanced a long time ago by Charles Darwin, R.A. Fisher, and others since
3. We’re wishy-washy DiNOs
There was a time when conventional wisdom held that molecules don’t really matter, and that the real issues in evolution are about “phenotypes”, meaning gross morphology, organismal behavior, life history and so on. Selection is what matters, it was said, and selection acts on these things. Back then, studies of evolutionary processes that relied on molecular data alone were considered incomplete or unimportant.
IMHO, this was little more than prejudice. “Genotype” is an informational concept. Everything else is “phenotype”, including the chemical structures of genes, proteins, complexes, and so on. The disparaging view that the “molecular level” provides only a superficial view on evolution was developed and actively promoted by Mayr, Simpson, Dobzhansky and others in the 1960s and 1970s precisely to hide the utter failure of their own theory to anticipate the findings of molecular sequence comparisons, which quickly led to a new field of “molecular evolution” with its own journals and meetings. If the “Modern Synthesis” had actually anticipated molecular evolution, this wouldn’t have happened.
The notion that we can’t apply the Darwinian creativity-of-selection theory to molecules because Darwin didn’t know about molecules is lame. It’s somehow unfair to expect a theory of evolution to address the evolution of things the original author hasn’t seen yet? Forterre is defending Darwinism by shifting the goal-posts and making it superficial. He doesn’t actually believe in Darwinism, only in the Darwin Brand.
The value of this brand is sustained by way of false conceptions of history that allow the neo-Darwinian in-group to appropriate other views using flexible language, e.g., by redefining “gradualism” to include saltationism. How does this happen? Inside the bubble of neo-Darwinism, acolytes learn that all historic alternatives are various forms of idiocy. So, when faced with evidence that Mayr, et al. weren’t right, they will never conclude that Mayr, et al. were wrong, but will react with something like “well, they may have missed some things, but at least they were on the right side of the debate– they weren’t saltationists!”.
Yet there is no doubt that the historic gradualism debate pitted Darwinians defending the “all small” position against everyone else arguing for the “some non-small” position. Darwin said that nature does not take leaps, natura non facit salta. What could be clearer? As explained above, the “all small” position underlies the Darwinian position on the creativity of selection: it is a logical piece that fits together with other pieces in a self-consistent theory. It is not dispensible. This is why Darwin said that his theory would “utterly break down” if it could be shown that some complex organ had not arisen by numerous successive slight modifications. He literally used the word “infinitesimal” (also “slight” and “insensible”) to describe variations used by “natural selection.” As a concrete example of what he meant, Darwin once invoked a difference of 1/100th of an inch in the length of a bird’s beak, a difference so small it would be difficult to measure reliably. 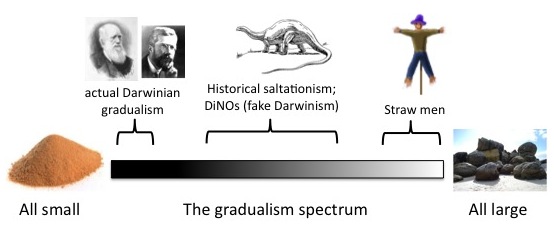
The logical alternative to “all small” is “not all small”, i.e., “some non-small.” Huxley objected to Darwin’s view because he believed that some changes take place by steps or jumps, not because he insisted that all changes take place by jumps.[4]
The same was true of the early geneticists who are ridiculed as “mutationists” or “saltationists”. They did not reject the idea of evolution via small-effect variations: they simply insisted that we need to incorporate discrete mutation-defined steps in our thinking. The idea that evolutionary change could happen relatively smoothly in a Mendelian world was not invented by Darwinians: Bateson outlined the multifactorial model back in 1902, when little Ronny Fisher was a schoolboy. Likewise, Johannsen was doing actual selection experiments with superficially smooth distributions of discrete variants long before Fisher’s 1918 paper that, in the self-serving neo-Darwinian view of history, proves that selection and Mendelism are compatible. Even de Vries, the most extreme advocate of macromutations among the early geneticists, accepted that gradual changes happen in evolution (for the views of early geneticists, see Stoltzfus and Cable, 2014).
Today, mainstream sources present Darwinian “gradualism” as a middle ground. When neo-Darwinians refer to saltationism, this is now a straw-man position in which all evolutionary changes are dramatic leaps, or in which major taxon-defining traits must appear in a single step (e.g., Coyne; but see [3]). For instance, note how the issue is framed by researchers cited by Chouard:
Many researchers have welcomed the return to favour of large-effect mutations and have resurrected Goldschmidt’s long reviled idea of the hopeful monster. But they can’t ignore the small-effect mutations. “We need much more data before the issue of large versus small can be settled”, says Coyne. Kingsley, like Coyne favours a middle-ground view, in which neither large- nor small-effect mutations are ruled out. “Our work has too often been portrayed as saying that Darwin was wrong” about big leaps in adaptation, he says. But in fact, none of the traits his group has studied is completely due to the effects of a single gene.
Actually, Kingsley’s work does show that Darwin was wrong. With the “single gene” comment, Coyne and Chouard are setting up a strawman position.
What is going on here? What is going on here is the phenomenon of Darwinism-in-Name-Only. Weldon was ready to die for Darwinian gradualism. By contrast, DiNOs aren’t committed to defending Darwin’s actual beliefs, which they do not share. In response to Shapiro’s criticism that molecular saltations speak against Darwinism, Dean (2012. Evolution: a View from the 21st Century. Microbe Magazine) objects thus:
His stance is patently unfair. Thomas Huxley famously criticized Darwin for championing too gradualist a view of phenotypic evolution. Today’s Darwinists accept Huxley’s criticism . . . Horizontal gene transfer, symbiotic genome fusions, massive genome restructuring (to remarkably little phenotypic effect in day lilies and muntjac deer), and dramatic phenotypic changes based on only a few amino acid replacements are just some of the supposedly non-Darwinian phenomena routinely studied by Darwinists.
That is “today’s Darwinists” don’t agree with Darwin, but with saltationists, but they still get to call themselves Darwinian gradualists. How does that work? Read Dean’s last sentence to understand the wishy-washy thinking behind this. Dean is implying that if someone who wears the Darwin label studies horizontal gene transfer, then the Darwin label automatically transfers to that phenomenon, regardless of whether it is consistent with Darwin’s stated views. [4] Think about it.
- Person P is sympathetic to theory T.
- Person P studies phenomenon X.
- Therefore, phenomenon X is part of theory T.
Obviously we are not talking about scientific theories anymore. Dean’s Darwinism is a constantly shifting pastiche of people and their interests and opinions.
It’s a form of identity politics. DiNOs identify, not with Darwin’s theory of evolution, but with The Darwin Brand and with historical predecessors who wore the Darwin Brand. The brand has power, including the power to appropriate credit. The brand has logos that are instantly recognizable to geeky scientists. Labeling an idea as “Darwinian” gives it automatic credibility with the community of self-styled “Darwinians”, and labeling an idea “non-” or “anti-Darwinian” automatically undercuts it [5] Defending the brand is a sacred obligation, like defending the family name. The brand has so much cred that scientists— normally desperate to make claims of novelty— are eager to align themselves with someone who has been dead for generations.
What the brand signifies is negotiable, subject to the constraints of history— a weak constraint, as language is flexible and little history is known to the brand’s defenders [6]. DiNOs recoil at the word “saltationism,” not because they object to the idea that there are discrete steps in evolution (i.e., what saltationism actually means), but because it offends them culturally— the Darwin Brand suffers whenever someone utters the phrase “arrival of the fittest” (de Vries) or invokes “saltation” seriously.
Are we saltationists yet?
This blog has gotten too long (as do most of them, alas), so I will wrap it up rather clumsily and inconclusively.
Based on evidence from molecular evolution, evo-devo, comparative genomics, and quantitative genetics, we have accepted the fact of saltationism: evolution sometimes takes jumps or steps. The variations that are incorporated in evolution are not entirely slight or infinitesimal, as Darwin believed under the theory he called “natural selection” (today, the words “natural selection” mean something different). Large changes, contrary to Darwin’s belief, are not always prohibitively deleterious. The incipient stages of useful structures, contrary to Darwin’s belief, are not always infinitesimal shifts.
But we are not saltationists yet. Partly this is because of the influence of conformists who continue to imply (without being entirely clear) that the body of evidence for saltation is not extensive enough to pass some intuitive threshold of importance. They believe that we have to do more work to know whether saltations are important enough to matter. They don’t feel any obligation to re-think the role of variation in evolution, which for them is still a matter of “raw materials”.
That’s one reason we haven’t fully embraced the implications of saltationism. Another reason is that, even for people who want to re-think the role of variation in evolution, it has proven difficult (e.g., consider Shapiro’s book as a failed attempt, recasting variation as “natural genetic engineering”). Perhaps I will say something more about that at another time. The fact that so many reformers (e.g., Andreas Wagner) invoke “the arrival of the fittest”— the de Vriesian catch-phrase from the saltationism debate 100 years ago— is promising. We may be in the midst of a minor renaissance of interest in the role of variation in evolution, something that I suggest in The Revolt of the Clay.
At the same time, I’m deeply skeptical about the potential for reform in evolutionary biology. Conformists have established the precedent that they can shift the goal-posts at will, distort history, change the meaning of words, and appropriate anything— even the Neutral Theory— in the name of neo-Darwinism and the Modern Synthesis. And reformers, for reasons that I cannot comprehend, have acquiesced to this. As a result, they cannot point out that the historic Modern Synthesis is false (which it is), or that there are any new causes (which there are), because they have agreed that DiNOs can shift the meaning of “Modern Synthesis” to keep it up to date, and they have agreed with the DiNOs that there can be no such thing as a competing causal theories in biology, but only one unified school of thought including a range of opinion that either is, or is not, sufficient. So the reformers have to convince us that their “extended synthesis” is unified (it isn’t) and that we should accept it because we aren’t already thinking broadly enough, because we need some “alternative narratives”. If evolutionary biologists worked in sociology departments, they might be more welcoming of “alternative narratives”. The current reform movement has given up all their strategic advantages and is going nowhere with arguments like this.
Notes
[1] Note that Darwin came to this position, not by reasoning upwards from known mechanisms, but by reasoning downwards from a high-level generalization. He began with the belief that organisms are exquisitely adapted, down to the finest detail. The evolutionary process that dominates in nature, he reasoned, must be the kind of process that allows for organisms to be exquisitely shaped by external conditions— a process based on an abundance of infinitesimal effects that can be pushed in any direction. Organisms could not be so exquisitely adapted if evolutionary change depends on distinctive individual variants. BTW, I’m not simply imagining thoughts in dead Darwin’s head— textual evidence indicates that Darwin reasoned backwards from adaptationism to gradualism, e.g., in his 1860 letter to Asa Gray in which he writes:
I reflected much on the chance of favorable monstrosities (i.e., great and sudden variations) arising. I have, of course, no objection to this, indeed it would be a great aid, but I did not allude [in OOS] to the subject for, after much labor, I could find nothing which satisfied me of the probability of such occurrences. There seems to me in almost every case too much, too complex, and too beautiful adaptation, in every structure, to believe in its sudden production.
Darwin nearly always embellishes his case against discrete changes by referring to them with dramatic language as “monstrosities”. Describing saltations in provocative and negative language is a common rhetorical trick of Darwinians, e.g., Wright associates them literally with “miracles”:
“From assisting Prof. Castle, I learned at firsthand the efficacy of mass selection in changing permanently a character subject merely to quantitative variability. Because of this and a distaste for miracles in science, I started with full acceptance of Darwin’s contention that evolution depends mainly on quantitative variability rather than on favorable major mutations. ” (Wright S. 1978. The Relation Of Livestock Breeding To Theories Of Evolution. Journal Of Animal Science 46:1192-200.)
The annoying tendency of Darwinians to depict themselves as standing above other less rational scientists, and to depict alternatives as magical or irrational, has the even more annoying consequence of making it difficult to determine what they really think. Clearly Darwinians are against monstrosities, but how large of a non-monstrosity will they tolerate? Clearly Wright is against miracles and for gradualism, but what exactly does that mean? When gradualism seems to fail, will he claim that there are no miracles and insist he was right all along? One way to get around this problem is to consider actual concrete cases. For instance, Darwin refers to the Ancon sheep as a “monstrosity”. Other scientists like Huxley did not describe it as monstrous. The Ancon sheep clearly arose in a single step, and along with the short legs that made it easier to contain in fenced areas, it also had undesirable phenotypes (such as joint problems) that presumably contributed to its eventual demise.
[2] MA studies attempt to characterize the spectrum of new variation. Variation was not ignored previously, but it was studied in a different way. In particular, the focus was on standing variation. Modern Synthesis population geneticists were obsessed with variation, but this was not a burning desire to understand how the introduction of novelty by mutation shapes evolution. Instead, what Gillespie refers to as The Great Obsession was to understand “the maintenance of variation in the gene pool”. This refers to a slightly different flavor of the argument for the creativity of selection. The classic argument is that selection is creative because any significant change is the accumulation of countless infinitesimal variations. Once Mendelism was established and it was clear evolution by fixation of distinctive mutants was a real possibility, Darwin’s followers took a different tack: selection never waits for a new mutation, but finds all the needed variation (to respond to new conditions) in the “gene pool”, which soaks up and “maintains” variation; individual alleles have no constant meaning due to interactions; evolution is an inherently interactive, polygenic process of shifting gene frequencies from one multi-locus equilibrium to another. In the works of Mayr, Dobzhansky, et al., this was largely a hand-waving argument. As Provine noted bitterly in 2001, the “gene pool” turned out to be one of the most superficial concepts in population genetics. However, the goal of understanding “the maintenance of variation in the gene pool” drove a population-genetics research program for a generation after the Modern Synthesis was established.
[3] In his response to the NYT blog by Olivia Judson, Coyne defines “macromutations” and “saltations” as referring to a position in which “mutations produce, in one fell swoop, complete new features that distinguish major groups, such as the extra pair of wings in dragonflies, the jaws of vertebrates, or the feathers of proto-birds.” Certainly many early geneticists suggested that new features could arise suddenly, in a definite form, often citing actual examples of mutants proving that new features can arise suddenly (and these suggestions play a prominent role in the writings of de Vries or especially Goldschmidt). Also, many critics of Darwinism have suggested that the features that distinguish higher taxa are not the same as the features that typically distinguish species (I confess that I have never really understood this argument). To yoke these two ideas together and dub them “saltationism” is to create a straw-man. This is simply not what saltationism means. In fairness to Coyne, we must take into account that he is responding to Judson’s piece that focuses on the idea of “traits likely to have evolved in one jump, rather than gradually”. That is, both Coyne and Judson are focused more on the trait-appears-in-one-step idea than on saltationism per se. This was a foolish way for Judson to frame the issue. But Coyne responds by tarring everything with the same brush, even taking aim at Gould’s punctuated equilibrium as a form of saltationism, which is patently incorrect (in Gould’s theory, the punctuations are merely rapid, not necessarily composed of non-infinitesimal increments). The second piece, Chouard’s Nature piece, is more focused on the idea of non-infinitesimal or large-effect changes, and less focused on the trait-appears-in-one-step idea.
[4] What was the nature of Huxley’s saltationism? In his review of OOS, Huxley objected to Darwin’s natura non facit saltum, saying that “nature does make jumps now and then, and a recognition of the fact is of no small importance in dispensing with several minor objections to the doctrine of transmutation.” In a letter to Bateson, Huxley wrote approvingly that “I see you are inclined to advocate the possibility of considerable ‘saltus’ on the part of Dame Nature in her variations. I always took the same view, much to Mr. Darwin’s disgust, and we used often to debate it. ” Thus, Huxley and Darwin had a genuine disagreement over whether evolution does not take jumps (Darwin), or takes jumps “now and then”, i.e., occasionally (Huxley). Note that, in spite of aligning himself with Bateson’s position, not Darwin’s, Huxley is sometimes called a “Darwinian” in the 19th-century sense of someone who stood with Darwin against the creationists. Those early “Darwinians” had diverse views of how evolution worked, including views that Darwin himself clearly rejected— which is why historian Peter Bowler referred to the initial success of evolutionism as “the non-Darwinian revolution”. Subsequently, Darwin’s most faithful followers took up the intra-scientific battle against saltationism, Lamarckism, and orthogenesis. That is, after “Darwinians” won the extra-scientific battle with creationism in the 19th century, there was an intra-scientific battle that distinguished “Darwinism” from other views of evolution. T.H. Huxley was a Darwinian in the first battle, but a non-Darwinian by the standards of the second battle.
[5] Many examples could be given of how hysterically evolutionary biologists react to the label “non-Darwinian”. The 1969 proposal of the neutral theory by King and Jukes was entitled “Non-Darwinian Evolution”. This title is universally described as being provocative, even though the theory they described was something completely foreign to Darwin’s thinking.
[6] Those who can put the Darwin label on their products command a higher price. Not having the Darwin brand is a liability. This has been true for a long time. Accordingly, there has been a struggle throughout the history of evolutionary biology to appropriate Darwin’s name. De Vries, for instance, argued that his view was the culmination and realization of Darwin’s thinking. I do not doubt de Vries’s integrity— he had a genuine admiration for Darwin that bordered on hero-worship. The point is that de Vries worked hard, rhetorically, to establish a fit between his view and Darwin’s, and we can understand this in economic terms as an expenditure of effort to obtain something valuable, capitalizing on the brand’s value. As related by Gould, 2002, others tried to prevent de Vries from appropriating the Darwin brand. Marsha Richmond’s 2006 piece (Isis 97:447–484) on the 1909 centennial and its ramifications notes the following:
In his review of Darwinism and Modern Science and Fifty Years of Darwinism [2 centenary volumes], the American entomologist Vernon Lyman Kellogg, author of the much-discussed recent work Darwinism To-Day (1907), recognized a key strategy employed by most contributors. “Each speaker,” he noted, “first recognizes the lasting debt of science and of himself to the immortal master, and then gently or forcfully [sic] proceeds to show how the master’s explanation fails to explain, and to unload and display his own precious personal baggage of ‘factors’; factors of environment, of mutation, of adaptation, of determinate variation, of isolation and what not.”41 Indeed, this was the general formula: authors traced their views back to Darwin so as to establish their legitimacy, then proceeded to outline their own special interpretations and conclude that Darwin would no doubt have agreed with them had he only known of this work. Even those who had been espousing their particular points of view for decades adopted this tactic.
This “legacy” trope is still played today, and often substitutes for a genuine scientific discussion.
Veiko Krauss
April 16, 2015 - 1:49 pm
Arlin,
I appreciate Your line of arguments. I would add that it is commonly accepted that novel genes arrive typically by duplications, hybridizations, horizontal gene transfers or endosymbiosis and only exceptionally de novo. At the same time, processes of sub- and neofunctionalization of gene duplications include necessarily nucleotide substitutions. Both will involved in nearly every phenotypically remarkable step of evolution.
To use a picture: A building constitutes of bricks and finery.
harry pinxteren
April 19, 2015 - 7:14 pm
arlin
great story. There’s one, little, poignant detail though. The argument with the Duke of Argyll was about a differenceo of an 1100th of an inch, not 1/100th !
http://www.darwinproject.ac.uk/letter/entry-4752
Arlin Stoltzfus
April 20, 2015 - 12:11 pm
Harry, I’m impressed that you caught the obscure reference! I’m going to guess that “1100th” in the URL you gave is a transcription error. The value is transcribed as “1-100th” in Poulton [1] which you can read online here:
http://books.google.com/books?id=omQCBvwUw1AC&pg=PA8&source=gbs_toc_r&cad=4#v=snippet&q=argyll&f=false
Ultimately one would have to see a facsimile of the letter to resolve this.
[1] Poulton EB. 1909. Fifty Years of Darwinism. In. Fifty Years of Darwinism: Modern Aspects of Evolution. New York: Henry Holt and Company. p. 8-56.
harry pinxteren
April 21, 2015 - 8:57 am
Arlin
I am impressed by your criminal investigation!
Anyway, 1100th or 1-100th, never mind, Poulton may have read the original letter, and the Darwin project may have it all wrong, these days we measure ‘new births’ not in inches but nm’s ( “in the sense in which the term wd usually be understood” )
I’ve been told that if you add juist one amino acid – I don’t know which one, but that doesn’t matter for the argument- to this one opsine a placozoa has, it becomes light sensitive, it can ‘catch photons’. Placozoa are blind as you know.
So, here we have some ‘new birth’ , I’d say. In about, 50 or 100 nm’s. Or even less? Anyway, you can’t have it very much smaller, can you?
If it’s true this example might become ‘paradigmatic’ as people in the US sometimes call it. So It might indeed make the difference between dark and light 😉