The most recent issue of PNAS includes a report by Galen, et al linking enhanced mutation at a CpG site to altitude adaptation in Andean house wrens (Troglodytes aedon), based on clear biogeographic and biochemical evidence of adaptation. I’ve been waiting for this, both in the narrow sense that I’ve been waiting for this particular study to appear in print, and also in the broader sense that I have been waiting for any paper on mutation-biased adaptation to appear in a prominent venue. Results like these, one hopes, will overturn the “raw materials” doctrine of neo-Darwinism and stimulate the development of a new understanding of the role of mutation in evolution.
Some parts of this story need further work, as suggested in the PNAS commentary.
However, I want to discuss broader implications, rather than dwell on uncertainties. So, I’m going to assume that Galen, et al are right, and that we see the Ile55 change in high-altitude wrens partly because it is beneficial, and partly because its occurrence is favored by a roughly 10-fold higher rate of mutation. That is, the higher mutation rate probabilized this particular adaptive change.
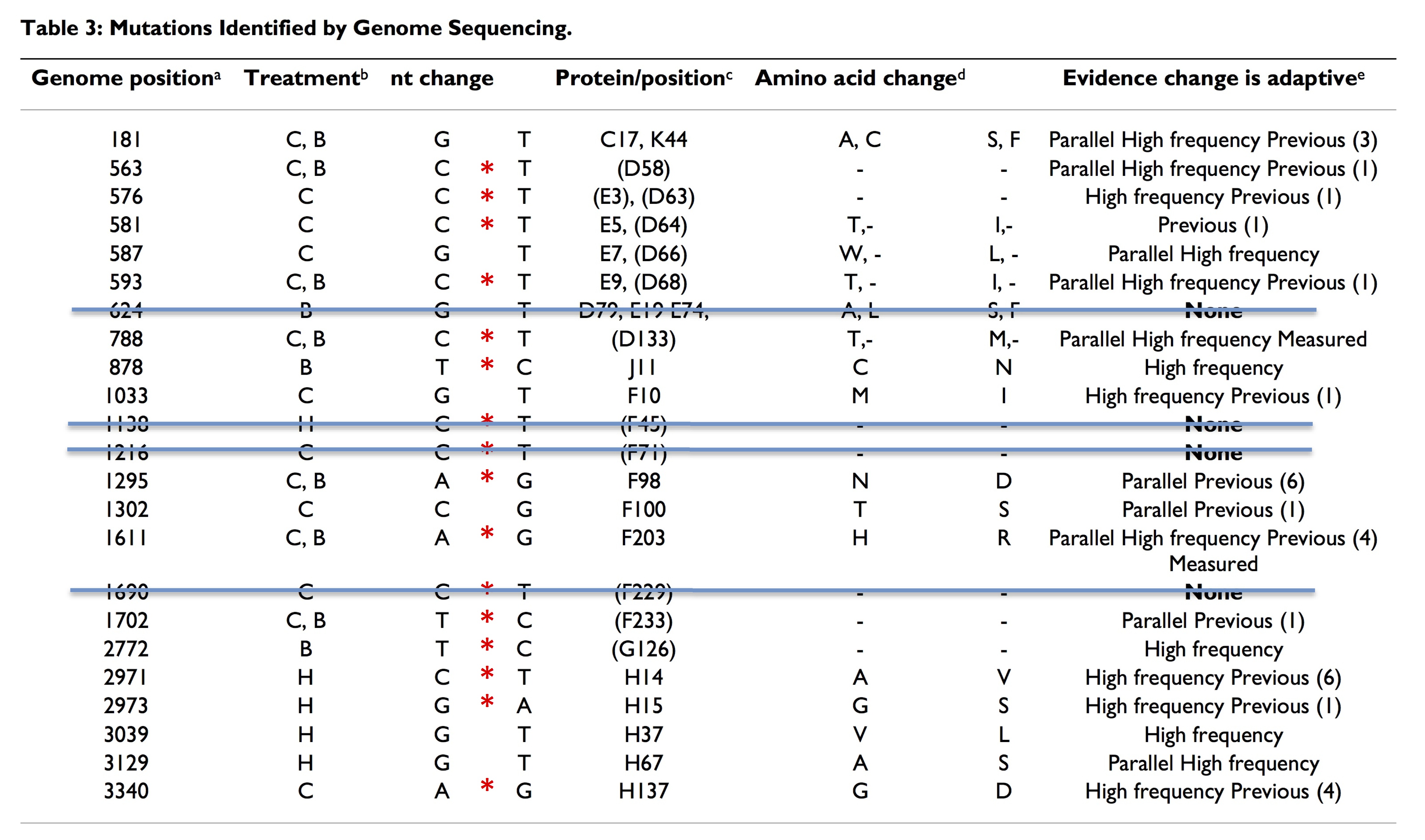
Furthermore, I’m going to assume that this is just the tip of the iceberg. If we look at other cases, we’ll see more evidence for a role of CpG hotspots. We’ll find that this effect applies to other types of mutation biases, e.g., transition-transversion bias, insertion-deletion bias, and so on. Furthermore, we will find that, when objective measures of effect-size are applied, the effects of mutational bias will be substantial. When I make this suggestion, it is not idle speculation. I’ve been tracking the evidence for years, and I reviewed some of it previously in The Revolt of the Clay and a followup (this is a more extensive list than appears in the PNAS commentary). For instance, in a study that I neglected to mention previously, Pepin and Wichman (2008) carry out repeated adaptation of phiX174, and the results show a clear bias toward transitions (above Table).
Excavating the memory hole
If all that is true, the implications for evolutionary theory are staggering. If “staggering” sounds like an exaggeration, a brief history lesson should set the record straight. Let’s review what leading authorities said throughout the latter half of the 20th century about the role of mutation and the influence of mutation rates (sources):
“The large number of variants arising in each generation by mutation represents only a small fraction of the total amount of genetic variability present in natural populations. … It follows that rates of evolution are not likely to be closely correlated with rates of mutation . . . Even if mutation rates would increase by a factor of 10, newly induced mutations would represent only a very small fraction of the variation present at any one time in populations of outcrossing, sexually reproducing organisms.” (Dobzhansky, et al., 1977, p. 72)
“mutations are rarely if ever the direct source of variation upon which evolutionary change is based. Instead, they replenish the supply of variability in the gene pool which is constantly being reduced by selective elimination of unfavorable variants. Because in any one generation the amount of variation contributed to a population by mutation is tiny compared to that brought about by recombination of pre-existing genetic differences, even a doubling or trebling of the mutation rate will have very little effect upon the amount of genetic variability available to the action of natural selection. Consequently, we should not expect to find any relationship between rate of mutation and rate of evolution. There is no evidence that such a relationship exists.” (my emphasis) (Stebbins, 1966, p. 29)
“Those authors who thought that mutations alone supplied the variability on which selection can act, often called natural selection a chance theory. They said that evolution had to wait for the lucky accident of a favorable mutation before natural selection could become active. This is now known to be completely wrong. Recombination provides in every generation abundant variation on which the selection of the relatively better adapted members of a population can work.” (Mayr, 1994, p. 38)
“The process of mutation supplies the raw materials of evolution, but the tempo of evolution is determined at the populational levels, by natural selection in conjunction with the ecology and the reproductive biology of the group of organisms” (Dobzhansky, 1955, p. 282)
“It is most important to clear up first some misconceptions still held by a few, not familiar with modern genetics: (1) Evolution is not primarily a genetic event. Mutation merely supplies the gene pool with genetic variation; it is selection that induces evolutionary change.” (Mayr, 1963, p. 613)
“if ever it could have been thought that mutation is important in the control of evolution, it is impossible to think so now; for not only do we observe it to be so rare that it cannot compete with the forces of selection but we know this must inevitably be so. “ (Ford, 1971, p. 361)
“Each unitary random variation is therefore of little consequence, and may be compared to random movements of molecules within a gas or liquid. Directional movements of air or water can be produced only by forces that act at a much broader level than the movements of individual molecules, e.g., differences in air pressure, which produce wind, or differences in slope, which produce stream currents. In an analogous fashion, the directional force of evolution, natural selection, acts on the basis of conditions existing at the broad level of the environment as it affects populations.” (Dobzhansky, et al., 1977, p. 6)
“Novelty does not arise because of unique mutations or other genetic changes that appear spontaneously and randomly in populations, regardless of their environment. Selection pressure for it is generated by the appearance of novel challenges presented by the environment and by the ability of certain populations to meet such challenges.”(Stebbins, 1982, p. 160)
According to this theory, every species has a “gene pool” that serves as a kind of dynamic buffer, soaking up and maintaining variation so that selection never has to wait for a new mutation. This buffer effectively insulates evolution from effects of mutation. As a result, mutations do not play a direct role in evolution, and they do not initiate change; rates of mutation are not determinative, so we don’t expect to see any correlation of the rate of evolution with the mutation rate. In general, mutation merely supplies raw materials, while selection is a higher cause, acting at a higher level to determine the direction and rate of change.
Near the end of this post, I will try to explain why the architects of the Modern Synthesis were so committed to a theory they could not have proved, and that seems hopelessly wrong today. For now, I just want to point out that this is a consistent position presented in forceful language, based on direct and confident appeals to concepts from population genetics.
A lopsided legacy
No one literally defends the “gene pool” theory anymore, nor does anyone (except Richard Dawkins) dismiss the role of mutation rates. Alas, this shift in mainstream beliefs was not accompanied by a revolution or any conscious restructuring of evolutionary theory. The reformist energy that should have gone into developing a mutationist alternative was sucked away by the Neutral Theory. Over time, Mayr and his cohort died off, and their intellectual descendants just stopped saying the things that were clearly wrong. Today we are left with a confused mixture, a Franken-theory with some zombie parts that just won’t go away.
Consider the neo-Darwinian catechism on the role of variation in evolution, which consists of 3 concepts. The first 2, which go back to Darwin’s time, are that mutation is merely a source of “raw materials” and merely a source of “chance.” The view of mutation and selection as opposing forces was developed by the Modern Synthesis. 
Apparently, it is not widely known that the raw materials doctrine refers to raw materials (e.g., crude oil, seawater, logs, coal, etc), and that it evokes Aristotle’s 4-fold classification of causes— “material” causes being the lowest kind. For instance, a shirt is made from fabric, fabric is woven from thread, and the thread is spun from either (1) natural fibers, in which case the raw materials are cotton, silkworm cocoons, etc, or (2) synthetic fibers, in which case the raw material is crude oil. Fabric is a material, but not a raw material. “Raw” materials are raw, unprocessed, unrefined.
Materials do not make the shirt happen, and do not dictate the size or shape. Some agent has to spin the thread, weave the fabric, cut it, and sew the pieces together. More generally, material causes are passive, and provide substance only, not form or initiative or direction. The final product is not implied or embodied in the materials. Instead, some active force or agent gives shape and form to the materials.
That is, the “raw materials” doctrine is a deliberate attempt to depict variation as a kind of passive clay that can be molded into anything, with selection as the agent— the active force shaping outcomes. In the Darwinian “gradualist” view, a single variation contributes to adaptation in the same way that a single grain of sand contributes to a sand-castle (see Why Size Matters: Saltation, Creativity and the Reign of the DiNOs). I suspect that if scientists stopped to consider what the “raw materials” doctrine intends, they would stop repeating it. For instance, one sees “raw materials” in the evo-devo literature, but no one in evo-devo actually believes this— they are all searching for developmental mutations that change body plans and reconfigure toolbox genes and generally make fantastic things happen.
The view depicting mutation and selection as opposing forces, with mutation too weak to overcome selection, arose from an early mathematical result called the mutation-selection balance, which represents how often we would expect to see a particular disease-causing mutation. This view makes mutation-biased adaptation seem like an impossibility. Below, I’ll use a population-genetics model to explain why this view misleads us.

Finally, if mutation is merely a source of “chance”, then how can a bias in mutation make evolution more predictable?
These old ways of thinking aren’t helpful. They don’t make the possibility of mutation-biased adaptation suggested by Galen, et al. comprehensible. We’ve inherited a lop-sided legacy. When 3 generations of scientists are taught that mutation merely supplies raw material, that it is a weak force, a source of chance, etc., we can’t expect this to promote a deep understanding of mutation. Like the character of Reggie in M. Night Shyamalan’s The Lady in the Water, we’re only flexing our intellectual muscles on one side
Re-thinking the role of mutation
To develop a new understanding of the role of mutation in evolution, let’s start by re-thinking the common metaphor of evolution as a climbing algorithm. Imagine, as an analogy for evolution, a climber operating on the jagged and forbidding landscape of Les Drus (Figure). A human climber would scout a path to a peak and plan accordingly, but a metaphor for evolution must disallow foresight and planning, therefore let us imagine a blind robotic climber. The climber will move by a two-step mechanism. In the “proposal” step, the robotic climber reaches out with one of its limbs to sample a point of leverage, some nearby hand-hold or foot-hold. Each time this happens, there is some probability of a second “acceptance” step, in which the climber commits to the point of leverage, shifting its center of mass.

Biasing the second step, such that relatively higher points of leverage have relatively higher probabilities of acceptance, causes the climber to ascend, resulting in a mechanism, not just for moving, but for climbing.
What happens if a bias is imposed on the proposal step? Imagine that the robotic climber (perhaps by virtue of longer or more active limbs on one side) samples more points on the left than on the right during the proposal step. Because the probability of proposal is greater on the left, the joint probability of proposal-and-acceptance is greater (on average), so the trajectory of the climber will be biased, not just upwards, but to the left as well. If the landscape is rough, the climber will tend to get stuck on a local peak that is upwards and to the left of its starting point.
Now, let us take this idea and make it into a population-genetics model, following Yampolsky and Stoltzfus (2001). 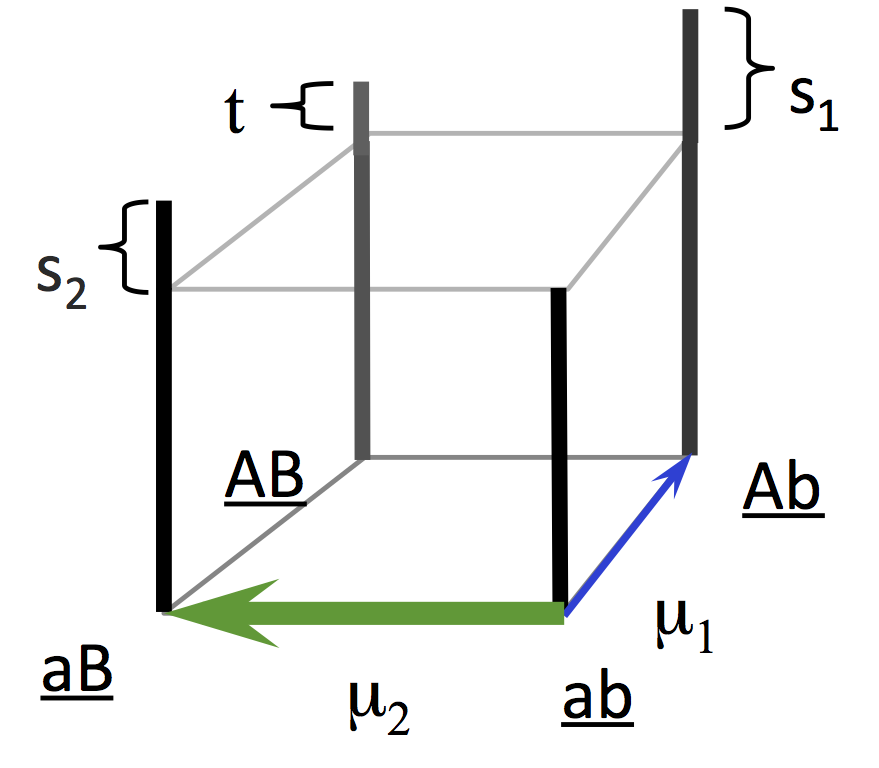
We’ll start with an ab population and evolve either to Ab or aB. That is, we are going on a one-step climb, and we’ll climb either to the left, or to the right. Obviously, if Ab is the more fit alternative (i.e., if s1 > s2) and the mutation rate to Ab is higher (u1 > u2), then Ab is favored. But what if Ab is the more fit alternative, and mutation favors aB? That’s the critical case.
The results are shown at right. 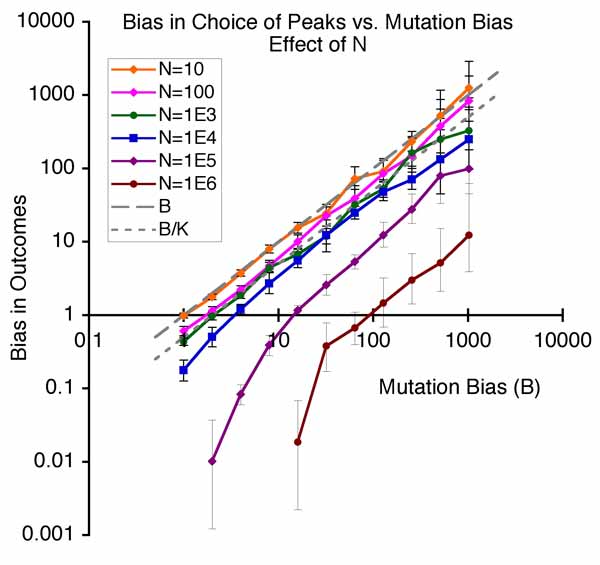 The upward slope indicates that the bias in outcomes (toward the mutationally favored peak) increases with increased mutation bias. In the smallest population, we are looking at neutral evolution, where only the bias in mutation (dashed line showing B = u1/u2) matters. As population size increases, we enter the regime of origin-fixation dynamics (or what Gillespie calls “strong selection, weak mutation”), where there is a proportional effect of both mutation bias and fixation bias, shown by the dotted line, which is (s1/s2)*(u1/u2)
The upward slope indicates that the bias in outcomes (toward the mutationally favored peak) increases with increased mutation bias. In the smallest population, we are looking at neutral evolution, where only the bias in mutation (dashed line showing B = u1/u2) matters. As population size increases, we enter the regime of origin-fixation dynamics (or what Gillespie calls “strong selection, weak mutation”), where there is a proportional effect of both mutation bias and fixation bias, shown by the dotted line, which is (s1/s2)*(u1/u2)
As population size gets larger and uN is no longer small, we depart from strict origin-fixation dynamics, but there is still an effect of mutation bias (uN is about 1 and 10 for the 2 largest populations).
Evolution doesn’t have to work this way. In a previous post I invoked 2 different styles of self-service restaurant— the Buffet and the Sushi Conveyor— to compare and contrast 2 different regimes of population genetics. The sushi conveyor offers a dynamic, iterated process of proposal and acceptance.

We choose (we select), but we don’t control what is offered or when: instead, we accept or reject each dish that passes by our table. This is like origin-fixation models of evolutionary dynamics, which depict evolution as a discrete 2-step process of mutational origination followed by fixation or loss (by selection or drift).

The architects of the Modern Synthesis viewed adaptation differently, according to the buffet model. Just as the staff who tend the buffet will keep it stocked with a variety of choices sufficient to satisfy every customer, the gene pool “maintains” abundant variation sufficient to meet any adaptive challenge (selection never has to wait for a new mutation). Adaptation happens when the customer gets hungry and proceeds to select a platter of food from the abundance of available choices.
A bias in variation will operate completely differently in the two models. Let us suppose that the buffet has 5 apple pies and 1 cherry pie. This quantitative bias in what is offered to the customer makes no difference. A customer who prefers cherry pie will choose a slice of cherry pie every time; and likewise a customer who prefers apple will choose apple. But at the sushi conveyor, the effect of a bias will be different. Let us suppose that occasionally a dish of sashimi comes by on the conveyor, with a 5 to 1 ratio of salmon to tuna. Even a customer who would prefer tuna in a side-by-side comparison may end up choosing salmon more frequently— a side-by-side comparison simply is not part of the process.
The reason I like this metaphor is that we can relate it directly to population genetics. The sushi conveyor corresponds to the origin-fixation regime in which the chance of making a choice is directly correlated with the mutation rate, because each change depends on a new mutation. By contrast, in the “gene pool” (buffet) regime, all the variants relevant to the outcome of evolution are present initially. Using the model above, if we put the alternative genotypes aB and Ab into the starting population at just 0.5 % frequency, this kills the effect of mutation bias completely, as shown by the flat lines in the figure below.
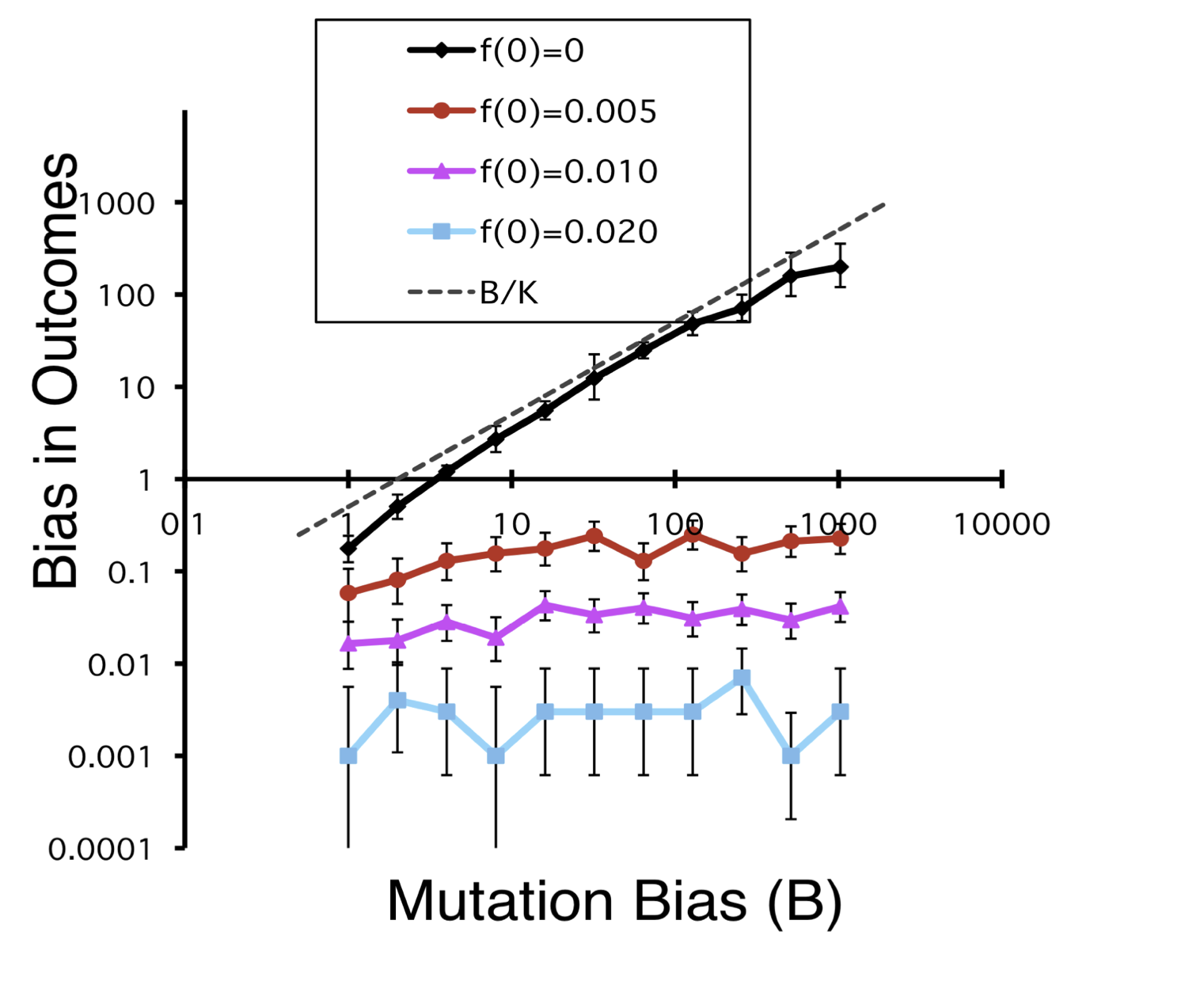
Why is selection so much more effectual in the buffet regime? The probability of fixation of a new beneficial mutation is about 2s. If sAb = 0.02 and saB = 0.01, this 2-fold difference in s corresponds to a 2-fold preference for Ab in the origin-fixation regime. In the buffet regime, the impact of exactly the same fitness difference is far greater: if we have 2 alternative alleles already in a population and they have escaped the drift barrier, selection pretty much always establishes the more fit alternative.
Why is mutation bias so important in one regime but not in the other? The bias operating so effectively in the origin-fixation regime is a bias in the introduction process, i.e., a bias in the rate of introduction of new alleles. This kind of bias is a profoundly important effect that directly impacts the course of evolution. But in the buffet regime, there is no bias in the introduction process, because there is no introduction process— all relevant alleles are present already. Once the alleles are present, mutation can shift their relative proportions in a biased way, but such shifts are quantitatively trivial compared to the shifts caused by selection (or even drift). This is why the architects of the Modern Synthesis said that mutation is a “weak force”.

Let’s return to metaphors one last time. For a century, we have understood that mutation and selection are both necessary. But the role of mutation has been depicted as supplying raw materials or chance. When seen as a force, mutation is said to be weak. We have imagined selection in charge without any mutational effects, or mutation in charge when selection is absent (neutral evolution), but making them co-pilots is a new way of thinking (Figure).
How the Modern Synthesis got population genetics wrong
I promised earlier that I would explain why the architects of the Modern Synthesis were so committed to a view that they had not proved, and which led them to believe that the rate of evolution would not reflect the rate of mutation.

First, let’s review the classic consensus on the genetic basis of evolutionary change. Prior to the molecular revolution, the Modern Synthesis held that change consists of a smooth shift, in the interior of a gene-frequency space, from the previous optimal multi-locus distribution of allele frequencies to the new optimum. The “shifting gene frequencies” consensus equated evolution with adaptation, and stressed that evolutionary change is
- initiated by an environmental shift (which disrupts the current optimum)
- driven by selection
- fueled by available variation + recombination (“gene pool”)
- not dependent on new mutations
- multi-factorial, involving many loci each with small effects
Why were the architects of the Modern Synthesis so strongly committed to an elaborate view that they hadn’t proved? Welcome to the world of science. In science, there are theories, conceptual systems for generating explanations and predictions through formal and informal reasoning (e.g., the metaphors and analogies used above). The predictions of theories like the Modern Synthesis are not always bottom-up predictions. If some high-level proposition Y is accepted as true, a lower-level proposition X is implied if X is the only way to get to Y. This is not the logical fallacy of affirming the consequent: if the theory asserts the truth of Y, and X is necessary for the truth of Y, then X (and of all of the implications of X) are predictions of the theory. We can put this in more flexible Bayesian terms to the effect that, if Y is more likely when X is true, then evidence for Y increases our belief in X.
In this case, what are X and Y? The Modern Synthesis was committed to the buffered “gene pool” view (X), because this is the particular view of population genetics that justifies Darwinian doctrines of gradualism, the creativity of selection, the subordinate status of variation as a source of random raw materials, and the control of selection over the direction of evolution (Y). Stripped of all nuance, the core claim of Darwinism is that, because organisms are exquisitely adapted, down to the finest detail, the mechanism of evolution must supply abundant infinitesimal raw materials for selection to shape the organism precisely to conditions. The Modern Synthesis “gene pool” view provides the needed mechanism. Alas, it’s mistaken.
Synopsis
Once again, I’ve said way too much, so let’s review the big picture.
For decades, a minority of evolutionists have been fascinated by the influence of biases in mutation in shaping genes, proteins and genomes. For decades, mainstream scientists have been dissecting natural cases of adaptation, and also carrying out adaptation in the lab. Those previously separate research directions come together in Galen, et al, and in some other studies such as Couce, et al., 2015 and Meyer, et al (see The Revolt of the Clay). The results suggest that the textbook doctrine on the role of mutation in evolution is incorrect:
- raw materials: mutation is not merely supplying raw materials because, in the case reported by Galen, et al, a single mutation changes affinity by 34 % (i.e., its not like 1 sand-grain among thousands)
- mere chance: mutation is not merely a source of chance because, in this case, the bias in mutation makes evolution more predictable, not less predictable
- weak force: evolution does not follow the logic of opposing forces where either selection or mutation must prevail, but instead allows simultaneous dual causation
For those who don’t care about theory, concepts, or history, the point of Galen, et al (and the other studies cited above) is that ignoring non-randomness in mutation means ignoring a potentially important source of information about what is likely to happen in evolution, and conversely, studying the rates of mutations gives us more leverage to predict and explain evolution.

Leave a comment